![]()
Эволюция ящериц и змей сопровождалась изменениями Hox-генов
|
Hox-гены играют ключевую роль в эмбриональном развитии животных, управляя дифференцировкой отделов тела вдоль передне-задней оси. Ученые из Швейцарии, Новой Зеландии и США показали, что разнообразие планов строения, наблюдаемое у чешуйчатых рептилий (ящериц и змей), связано со значительными изменениями в структуре и работе «задних» Hox-генов, которые у наземных позвоночных отвечают за формирование поясничного, крестцового и хвостового отделов. В окрестностях этих генов у чешуйчатых обнаружено много встроившихся мобильных генетических элементов, чего не наблюдается у других четвероногих.
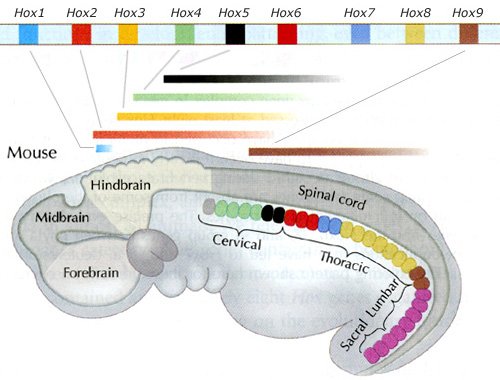
Важнейшая функция Hox-генов состоит в том, что они подробно «размечают» эмбрион вдоль передне-задней оси. Дальнейшая судьба эмбриональных клеток, оказавшихся в той или иной части эмбриона, зависит от набора Hox-генов, работающих (экспрессирующихся) в этой области. Для каждого Hox-гена характерна определенная область экспрессии. Например, гены Hox12 и Hox13, как правило, работают только в задней части эмбриона, которая в дальнейшем станет хвостом; гены Hox10 у некоторых позвоночных работают от заднего конца эмбриона до той черты, которая станет границей между грудным отделом (где на позвонках есть ребра) и поясничным, где ребра не развиваются, и т. д. «Hox-код», определяющий план строения организма, довольно сложен и не совсем одинаков у разных групп позвоночных. Вряд ли можно сомневаться в том, что многие крупные эволюционные преобразования, затрагивающие план строения, были связаны с изменениями в структуре и экспрессии Hox-генов. Однако хорошо изученных примеров, иллюстрирующих эту связь, пока немного (см., например, о связи утраты ребер на поясничных позвонках с генами Hox10: Новая палеонтологическая находка проливает свет на раннюю эволюцию млекопитающих, «Элементы», 17.03.2007).
У многих животных, в том числе у позвоночных, Hox-гены в геноме располагаются кластерами, то есть группами вплотную друг другу. Самое удивительное, что порядок расположения генов в Hox-кластерах часто (хотя и не всегда) совпадает с распределением областей экспрессии вдоль передне-задней оси: впереди находятся «головные» гены, за ними следуют гены, отвечающие за формирование средних участков тела, а замыкают кластер «задние» гены, управляющие развитием задних частей туловища (см.: Новое в науке о знаменитых Hox-генах, регуляторах развития, «Элементы», 10.10.2006).
|
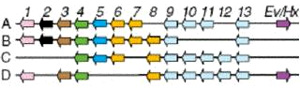
У предков позвоночных, как у современного ланцетника, в геноме был всего один Hox-кластер, включающий 14 генов. На ранних этапах эволюции позвоночных произошли две последовательные полногеномные дупликации (учетверение генома). В результате позвоночные приобрели четыре Hox-кластера вместо одного исходного. Это открыло перед позвоночными большие эволюционные возможности (см.: Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных, «Элементы», 23.06.2008). Отдельные Hox-гены в некоторых кластерах были утрачены, но в целом их набор и порядок расположения остался весьма сходным во всех четырех кластерах.
Паралогичные гены (то есть копии одного и того же Hox-гена в разных Hox-кластерах) во многих случаях приобрели немного различающиеся функции, что дало возможность весьма тонко регулировать эмбриональное развитие (см.: Многофункциональные гены — основа для эволюционных новшеств, «Элементы», 30.06.2008). Это сильно облегчило развитие новых планов строения в ходе эволюции позвоночных.
Группа исследователей из Швейцарии, Новой Зеландии и США детально изучила структуру и работу Hox-генов у чешуйчатых рептилий (отряд Squamata). Этот отряд, объединяющий ящериц и змей, интересен большим разнообразием планов строения и вариабельностью признаков, непосредственно связанных с передне-задней дифференцировкой туловища (относительная длина отделов тела, число позвонков в них и т. п.). Поэтому логично было предположить, что Hox-кластеры чешуйчатых должны обладать какими-то специфическими особенностями и что Hox-гены ящериц и змей должны чем-то различаться.
Ранее было показано, что области экспрессии «передних» Hox-генов у змей расширились в заднем направлении по сравнению с другими позвоночными. Это хорошо согласуется с общим удлинением тела. Кроме того, было установлено, что правило колинеарности (то есть одинаковый порядок расположения генов в кластере и областей их экспрессии в эмбрионе) у змей строго соблюдается. Авторы обсуждаемой работы сосредоточились на «задних» Hox-генах (от 10-го до 13-го).
|
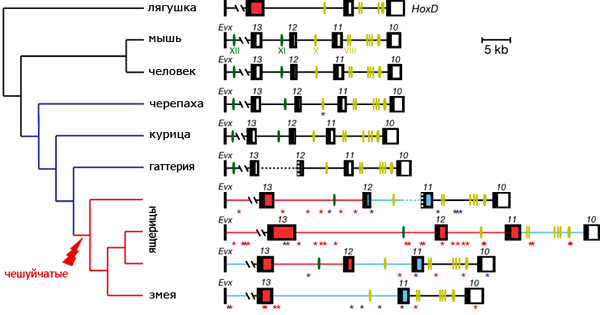
Самым неожиданным результатом оказалось присутствие в Hox-кластерах чешуйчатых большого количества встроившихся мобильных генетических элементов — транспозонов и ретротранспозонов, а также множества повторяющихся последовательностей. В результате общая длина задней части Hox-кластеров у чешуйчатых значительно выросла по сравнению с другими наземными позвоночными.
Всё это, по-видимому, говорит о том, что у чешуйчатых ослабли эволюционные ограничения, препятствующие накоплению изменений в задней части Hox-кластеров. «Очищающий» отбор, отбраковывающий подобные изменения у других позвоночных, в эволюции ящериц и змей явно действовал менее эффективно. Этот вывод подтвердился и в ходе анализа кодирующих участков Hox-генов. В этих участках у ящериц и особенно у змей, по сравнению с другими позвоночными, накопилось много значимых замен (то есть нуклеотидных замен, ведущих к изменению аминокислотной последовательности кодируемых белков). Одни из этих замен, по-видимому, зафиксировались случайно, из-за ослабления очищающего отбора, тогда как другие закрепились под действием положительного отбора, то есть были полезными.
Авторы изучили характер экспрессии задних Hox-генов у эмбрионов хлыстохвостой ящерицы и маисового полоза. При этом обнаружилось много интересных деталей, подтверждающих исходное предположение о том, что изменения плана строения в ходе эволюции чешуйчатых были самым тесным образом связаны с изменениями в работе задних Hox-генов.
|
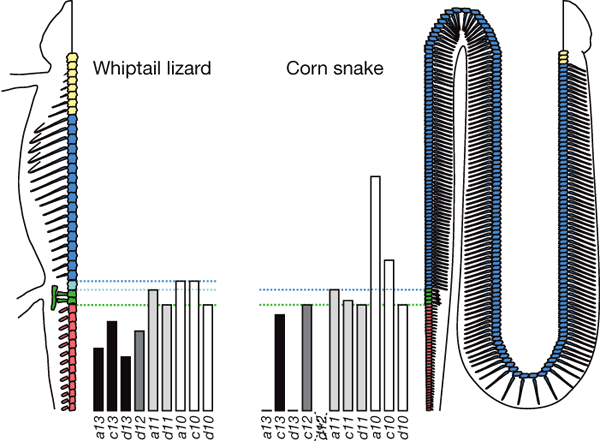
У ящерицы, как и у других наземных позвоночных, передний край области экспрессии генов Hoxa10 и Hoxc10 в точности соответствует границе между грудным и поясничным отделами. Одной из функций этих генов является подавление развития ребер. У змей нет поясничного отдела, а на бывших крестцовых позвонках (у змей они называются «клоакальными») имеются особые раздвоенные ребра. По-видимому, эти морфологические особенности связаны с тем, что гены Hoxa10 и Hoxc10 у предков змей утратили способность останавливать рост ребер. Область экспрессии этих генов у полоза заходит далеко в грудной отдел. Предполагается, что эти гены отвечают также за своевременное прекращение роста грудного отдела. По-видимому, эта их функция у змей тоже ослаблена, что могло быть одной из причин удлинения грудного отдела у змей по сравнению с их предками — ящерицами. Удлинение хвостового отдела у змей, по-видимому, связано с тем, что из четырех генов, «тормозящих» рост хвоста у ящериц (Hoxa13, Hoxc13, Hoxd13, Hoxd12) один ген у змей полностью утрачен (Hoxd12), а два других (Hoxa13, Hoxd13) не участвуют в передне-задней «разметке» эмбриона и используются только в формировании половых органов.
Обнаруженные факты, несомненно, помогут ученым продвинуться в расшифровке «Hox-кода», задающего план строения животных, и приблизиться к пониманию генетических основ макроэволюционных преобразований. Авторы отмечают, что многочисленные случаи независимой утраты или частичной редукции конечностей у чешуйчатых (см.: Утрата конечностей почти необратима, «Элементы», 10.12.2008) тоже могут быть связаны с тем, что в этом отряде рептилий задние Hox-гены получили нетипичную для других животных эволюционную «свободу». На них стал слабее действовать очищающий отбор, что позволило быстро накапливать разнообразные мутации.
Известно, что задние Hox-гены играют ключевую роль не только в оформлении задних отделов туловища, но и в развитии конечностей. Поэтому некоторые мутации этих генов, ведущие, например, к удлинению тела или к редукции поясничного отдела, теоретически могут приводить и к таким побочным эффектам, как редукция конечностей. Авторы также отмечают, что удлинение тела в сочетании с редукцией конечностей встречается и в других группах позвоночных (например, у некоторых амфибий). Было ли это связано с такими же изменениями в работе Hox-генов, как у змей, или с другими, покажут будущие исследования.
Источник: Nicolas Di-Poi, Juan I. Montoya-Burgos, Hilary Miller, Olivier Pourquie, Michel C. Milinkovitch, Denis Duboule. Changes in Hox genes’ structure and function during the evolution of the squamate body plan // Nature. 2010. V. 464. P. 99–103.
![]()