![]()
Чтобы превратить самок в гермафродитов, достаточно двух мутаций
|
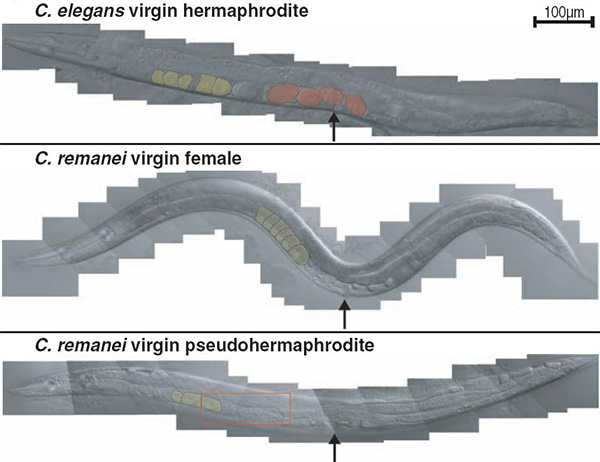
Большинство круглых червей рода Caenorhabditis — это обычные раздельнополые виды, состоящие из самцов и самок. Однако у двух видов этого рода (C. elegans и C. briggsae) самок нет, а есть самцы и гермафродиты, способные к самооплодотворению. Эксперименты на раздельнополых червях C. remanei показали, что для превращения самок в полноценных гермафродитов достаточно уменьшить активность двух генов (
1. Хорошая модель для изучения эволюционных новообразований
У большинства круглых червей (нематод), как и у многих других животных, пол определяется генетически, при помощи половых хромосом. Если в оплодотворенном яйце две
Однако у двух наиболее изученных видов нематод, Caenorhabditis elegans и C. briggsae, особи с двумя
Предки C. elegans и C. briggsae были нормальными раздельнополыми червями. Это следует, в частности, из того, что все остальные виды рода Caenorhabditis — раздельнополые. Таким образом, гермафродитизм у C. elegans и C. briggsae является эволюционно «молодым», новым признаком. Поскольку оба вида относятся к числу наиболее хорошо изученных животных, есть все основания надеяться, что в данном случае удастся расшифровать во всех деталях молекулярно-генетические механизмы становления этого нового признака в ходе эволюции.
2. Как сделать из самки гермафродита
Американские биологи из Медицинского университета в Стратфорде и Крейтонского университета сделали важный шаг в этом направлении, о чём сообщили в последнем номере журнала Science. Они экспериментально показали, что для превращения самок раздельнополого вида C. remanei в полноценных самооплодотворяющихся гермафродитов достаточно уменьшить активность всего лишь двух генов.
Ранее было установлено, что в определении пола у C. elegans важную роль играет ген
Это навело авторов на мысль, что если у самок раздельнополого вида C. remanei подавить активность гена tra-2, то самки, возможно, превратятся в гермафродитов. Начали они с того, что полностью отключили ген tra-2 при помощи РНК-интерференции. В результате из яиц с двумя X-хромосомами развились вместо самок вполне полноценные самцы, производящие нормальные спермии, но никаких яйцеклеток.
Тогда авторы справедливо рассудили, что они, очевидно, немного перестарались. Ведь у настоящих гермафродитов активность гена tra-2 хоть и снижена, но не до нуля. Когда эту ситуацию удалось воспроизвести у C. remanei, просто уменьшив силу воздействия на экспрессию tra-2, на свет появились особи, которых авторы назвали «псевдогермафродитами». Тело у них было «женское», но в гонадах формировались одновременно и яйцеклетки, и спермии. Правда, эти существа оказались неспособны к самооплодотворению (отсюда и приставка «псевдо»). После скрещивания с нормальным самцом превдогермафродиты откладывали оплодотворенные яйца, но в меньшем количестве, чем нормальные самки. Это значит, что яйцеклетки у них более или менее нормальные, а неспособность к самооплодотворению, скорее всего, объясняется дефектами спермиев.
Чтобы выяснить, в чём состоят эти дефекты, авторам пришлось немало потрудиться, проверяя одну за другой разные гипотетические возможности. Оказалось, что спермии псевдогермафродитов нормальны по всем параметрам, кроме одного: они неактивны, никуда не ползут и, в частности, не перебираются в сперматеки.
У нормальных нематод спермии активируются (начинают ползать) под воздействием белков, содержащихся в семенной жидкости. Сохранили ли спермии псевдогермафродитов способность активироваться под действием этих белков? Чтобы это проверить, авторы скрещивали псевдогермафродитов с самцами C. elegans. Самцы C. elegans охотно спариваются с самками C. remanei, но эти браки бесплодны (как-никак два вида разошлись, по имеющимся оценкам, около 80 млн лет назад). Однако после спаривания с самцами C. elegans псевдогермафродиты C. remanei откладывали оплодотворенные яйца, некоторые из которых оказались жизнеспособными, и из них вывелись нормальные самки C. remanei. Этот и другие эксперименты показали, что семенная жидкость C. elegans действительно способна активировать спермии псевдогермафродитов, и в результате самооплодотворение становится возможным.
Это открытие резко сузило круг потенциальных генов-мишеней, на которые нужно воздействовать, чтобы превратить псевдогермафродитов в полноценных гермафродитов. «Подходящим» геном оказался swm-1, который кодирует белок, подавляющий активность других белков — протеаз, активирующих спермии. Ген swm-1 отвечает за предотвращение преждевременной активации спермиев у самцов C. elegans, а также, по-видимому, выполняет ряд других функций. Авторы понизили активность этого гена у псевдогермафродитов (тоже при помощи РНК-интерференции) — и те сразу приобрели способность к самооплодотворению, к большой радости исследователей.
3. Пропасть можно перепрыгнуть в два прыжка, но для этого нужен мостик
Исследование показало, что для появления в ходе эволюции нематод рода Caenorhabditis нового признака — гермафродитизма — было достаточно всего лишь уменьшить активность двух генов, входящих в два разных регуляторных каскада. Для этого нужны две мутации. Однако одновременное возникновение двух мутаций, полезных только вместе, но не по отдельности, крайне маловероятно. Могли ли они появиться последовательно или это равносильно преодолению пропасти в два прыжка? Авторы рассматривают два возможных сценария данного эволюционного превращения:
1) Сначала произошла мутация, уменьшившая активность гена swm-1 у самок, что позволяет активировать собственные спермии (если они есть). Эта мутация поначалу была нейтральной, потому что самки еще не умели производить спермии. В дальнейшем возникла мутация, понизившая активность tra-2. Эта мутация сразу оказалась полезной (была поддержана отбором), потому что превратила самок в самодостаточных гермафродитов (о полезности гермафродитизма см. ниже). Но могла ли первая мутация не принести вреда, если известно, что ген swm-1, скорее всего, многофункционален? И вот тут-то на помощь приходит знание геномов изучаемых червей. Оказывается, в геномах раздельнополых видов имеется только одна копия гена swm-1, а у гермафродитных есть еще и вторая, немного отличающаяся копия. По-видимому, становлению гермафродитизма способствовала дупликация этого гена, что и позволило снять «адаптивный конфликт». Одна из копий изменилась, чтобы обеспечить своевременную активацию спермиев у гермафродитов, а вторая продолжила выполнение остальных функций (см.: Многофункциональные гены — основа для эволюционных новшеств, «Элементы», 30.06.2008).
2) Второй сценарий начинается с мутации, снизившей активность tra-2. Это привело к появлению псевдогермафродитов. Лишь после этого возникла и закрепилась мутация, снижающая активность swm-1. Однако псевдогермафродиты фактически являются всего лишь неполноценными самками: к самооплодотворению они не способны, а с ролью самок справляются хуже, чем настоящие самки. Поэтому на первый взгляд кажется, что отбор должен был сразу отсеять первую мутацию. Но вспомним, что самки C. remanei привлекательны для самцов других видов, а спаривание с этими самцами дает им возможность самооплодотвориться. Эта особенность могла стать «мостиком», по которому эволюционирующий вид сумел преодолеть опасный промежуточный этап и дождаться возникновения второй мутации. В некоторых ситуациях псевдогермафродиты, способные к самооплодотворению после спаривания с самцами других видов, могли иметь адаптивное преимущество (см. ниже).
Таким образом, «преодоление пропасти в два прыжка» при ближайшем рассмотрении оказывается не таким уж невероятным. Там всё-таки были мостики.
4. Изменение активности генов — самый «удобный» способ эволюционных преобразований
Изменение активности гена в сторону уменьшения или увеличения может произойти в результате самых разных мутаций. Теоретически, это могли быть мутации в регуляторных областях самих генов tra-2 и swm-1, или мутации генов-регуляторов, управляющих их работой, или мутации регуляторов регуляторов, и т. д. Важно, что в такой ситуации — когда оказывается выгодно уменьшить или увеличить экспрессию какого-либо гена — вероятность того, что случайная мутация, меняющая активность гена, окажется полезной, а не вредной, приближается к 1/2, то есть становится чрезвычайно высокой. Не случайно очень многие «прогрессивные» эволюционные преобразования, как теперь выясняется, были связаны с изменениями именно уровня активности генов, а не их белок-кодирующих последовательностей (см.: Эволюция человека сопровождалась изменением активности генов-регуляторов, «Элементы», 13.03.2006).
5. Проверьте, включена ли вилка в розетку
Многих раздражает этот «мудрый совет», неизменно присутствующий в инструкциях по устранению неисправностей в работе электроприборов. Однако за ним кроется универсальный принцип: во многих случаях нужного эффекта можно добиться, манипулируя только выключателями и не развинчивая весь механизм.
Конкретные мутации, ответственные за снижение активности генов tra-2 и swm-1 у предков C. elegans и C. briggsae, возможно, были разными. В частности, известно, что у гермафродитов первого вида в подавлении активности tra-2 участвует ген fog-2, отсутствующий у второго вида. Предки C. briggsae утратили ген fog-2, вероятно, за ненадобностью. У этого вида ключевую роль в обеспечении нормального сперматогенеза у гермафродитов играет другой ген — she-1. Этот пример показывает, что в ходе эволюции «переключатели» генетических регуляторных каскадов могут меняться, причем сами каскады — и их функции — остаются прежними. Это относится и к механизмам определения пола. Разделение на самцов и самок есть у большинства животных; соответственно, есть и генетические «программы» (большие и сложные) развития по мужскому и женскому пути. Однако ключевые переключатели, направляющие развитие по одному из двух путей, часто меняются в ходе эволюции. Например, у тех же нематод многие виды перешли от хромосомной детерминации пола к «средовой», то есть пол у них зависит не от генов, а от условий, в которых проходит развитие. В этом нет ничего чудесного: точно так же вы можете заменить электрический выключатель в своей комнате на сложную систему последовательно и параллельно соединенных тумблеров, кнопку или шнурок с кисточкой, не меняя при этом проводку и люстру. Управлять освещенностью, возможно, станет удобнее, но сам «фенотип» от этого не изменится: свет по-прежнему будет или включен, или выключен.
6. О пользе самооплодотворения
У читателей, помнящих нашу предыдущую заметку о нематодах (Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009), может возникнуть вопрос: зачем вообще кому-то понадобилось переходить к гермафродитизму, если эксперименты показали преимущество перекрестного оплодотворения над самооплодотворением? Дело в том, что преимущества гермафродитизма во многих случаях совершенно очевидны — именно поэтому и приходится ставить специальные эксперименты, когда хотят показать, что в каких-то других ситуациях перекрестное оплодотворение всё-таки лучше. Например, если участки, пригодные для жизни нематод данного вида, встречаются очень редко (это, по-видимому, вполне справедливо для представителей рода Caenorhabditis) и вероятность того, что хотя бы один червь попадет на новый, незаселенный участок, очень мала, то вероятность того, что на этот участок попадут сразу два червя — самец и самка — будет и вовсе ничтожной. Естественно, в такой ситуации преимущество получат черви, способные размножаться без посторонней помощи путем самооплодотворения.
Источник: Chris Baldi, Soochin Cho, Ronald E. Ellis. Mutations in Two Independent Pathways Are Sufficient to Create Hermaphroditic Nematodes // Science. 2009. V. 326. P. 1002–1005.
![]()