![]()
Транспозоны — это потомки гигантских вирусов, полагают канадские ученые
|

Канадские ученые открыли новые вирусы, которые в качестве мишени выбрали не клетки бактерий или эукариот, а других вирусов. Иными словами, это паразиты, заставляющие болеть паразитов. Подобных примеров пока известно только три, и все они обнаружены в последние несколько лет. Генетическая последовательность нового вируса оказалась во многом схожа с последовательностями гигантских транспозонов — особых мобильных элементов клетки. Это сходство дало основание предположить, что гигантские транспозоны произошли от «двойных» паразитов, подобных новому вирусу.
Транспозоны — подвижные элементы генома. Различают классы ретротранспозонов и ДНК-транспозонов, для которых характерны разные механизмы копирования. В свою очередь, ДНК-транспозоны подразделяются на те, что работают по принципу вырезать и вставить, есть кольцевые транспозоны, а есть и гигантские самореплицирующиеся транспозоны типа Maverick или Polinton (МП-транспозоны). Нуклеотидные последовательности МП-транспозонов составляют 9–22 тысяч пар нуклеотидов, в них закодировано около 20 белков. Этот набор относительно консервативен. В нём обязательно присутствует интеграза — фермент, отвечающий за встраивание вирусной последовательности в геном хозяина и работу с гетерохроматином. Интегразы характерны для ретровирусов. Кроме того, имеется аденозинтрифосфатаза, сходная с той, что имеется у ДНК-вирусов с двойной нитью; цистеинпротеаза, аналогичная аденовирусной. Есть еще один белок, по своей структуре сходный с основным капсидным белком фикоднавирусов (Phycodnaviridae), поражающих водоросли.
Как видно, генетический набор МП-транспозонов представляет собой смесь разных типов вирусов. Но тем не менее в качестве основной гипотезы обсуждается происхождение этих транспозонов от линии так называемых PRD1-вирусов, имеющих икосаэдрический (двадцатигранный) капсид и двойную нить ДНК. МП-транспозоны известны у широкого круга эукариотических организмов от простейших до рыб и птиц. Новое исследование касается именно этих транспозонов. Обнаружились новые удивительные данные, позволившие микробиологам Матиасу Фишеру и Куртису Шатлу из Университета Британской Колумбии (Канада) предложить смелую гипотезу о происхождении этих транспозонов.
В теплых морях в планктоне живет двужгутиковый Cafeteria roenbergensis. Он поражается гигантским вирусом CRoV, вызывающим лизис (разрушение) клеток. В клетках Cafeteria ученые нашли у вируса CRoV необычного соседа — мелкие темные кристаллы нанометровых размеров. Эти кристаллы, как выяснилось, тоже представители мира вирусов; новый вирус получил имя Mavirus (от Maverick Virus по названию транспозона). Найденный новый вирус не реплицируется без CRoV: если клетки Cafeteria заразить только Мавирусом, то лизиса не происходит и «лишние» РНК не продуцируются. Если же в клетке присутствует только CRoV без мавирусного сопровождения, то лизис клеток-хозев значительно увеличивается.
Получается, что этот вирус «мешает» CRoV размножаться, но сам по себе не обладает репликационной активностью. Это означает, что перед нами вирус, паразитирующий на вирусах, то есть использующий в качестве мишени вирусы. Такие уже известны ученым: один из них описан под именем Спутник (Sputnik). Он является паразитом гигантского мимивируса. Другой, найденный в антарктическом озере, был недавно описан и названный OLV (Organic Lake Virophage). Подобных вирусов предложили называть вирофагами (см. вирофаг Спутник), по аналогии с бактериофагами. «Элементы» уже обращались к этой теме (см.: Вирусы тоже болеют вирусными заболеваниями, 08.09.2008). Вирофаги зависят не столько от ферментной системы хозяйской клетки, сколько от ферментной машины приглянувшихся им вирусов. Ясно, что они не нуждаются в ядерных генах хозяйской клетки, как обычные вирусы.
Геном Мавируса представляет собой кольцевую ДНК, состоящую из примерно 19 тысяч пар нуклеотидов, 20 белок-кодирующих последовательностей, длина которых в среднем около 883 нуклеотида. Получается, что белок-кодирующие последовательности составляют около 93% всего генома Мавируса. Оставшийся фрагмент ДНК представляет собой устойчивый мотив стартовой последовательности, включающей считывание всей цепочки вирусных белков. Спутник и Мавирус имеют около 20% сходства в белок-кодирующей области, так что если связь между этими вирофагами и имеется, то очень древняя. Интегразы у них разные.
Существенно больше сходства обнаруживают последовательности Мавируса и МП-транспозонов. Четыре консервативных гена и три факультативных оказались общими у Мавируса и этих транспозонов. В частности, общий мотив (участок из примерно 180 нуклеотидов) обнаружился у интеграз. Ученые отметили также более менее схожую длину МП-транспозонов и Мавируса: 19 против 15 и 20. Кроме того, они обратили внимание на единообразное устройство ДНК-последовательностей в конечных участках Мавируса и МП-транспозонов.
Мало того, некоторые из генов Мавируса оказались гомологичны и генам слизевика Polysphondylium pallidum. Эти гены слизевика явно являются наследием транспозонного элемента. Нужно при этом заметить, что промотерного участка у этой вставки не нашлось. По-видимому, транспозон вставился в геном слизевика, утеряв промотер.
Столь множественные параллели между Мавирусом и МП-транспозонами позволили предположить, что они эволюционировали от общего предка. В этой дискуссии у канадских ученых оказались хорошие козыри. Логика подсказывает, что стоит обсуждать две версии. Первая: Мавирус — это сбежавший из клетки транспозон. Вторая: транспозоны — это освоившиеся внутри клетки потомки вирофагов. Если выбрать первый вариант, то придется признать, что наш вырвавшийся на свободу агент захватывал по пути гены многих хозяев, а затем непрошенно пристроился к другому вирусу. Кроме того, трудно понять, почему сбежавший агент всё же похож на Спутник и почему он оказался связан с другим вирусом. При этом нам придется допустить, что получившийся Мавирус приобрел промотерный участок независимо от других вирусов. Слишком много неувязок в этом детективном сюжете.
|
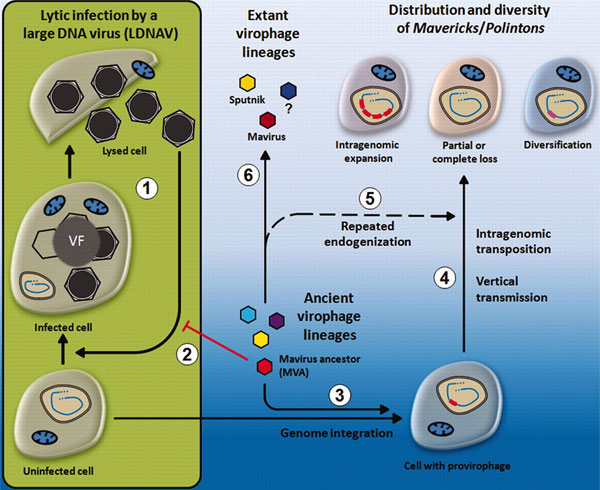
Их существенно меньше во втором сценарии. Согласно ему, у Мавируса и МП-транспозонов был общий предок. Он паразитировал на гигантском вирусе, внедрившемся в эукариотическую клетку. Тем не менее нужно учитывать, что вирофаг, этот вирусный прихлебатель, способствовал выживанию пораженной клетки, снижая лизис, повышая ее устойчивость к инфекции. Поэтому клетка поддерживала сожительство с вирофагом, оно было ей выгодно по принципу «враг моего врага — мой друг». Отбор способствовал широкому распространению вирофага в клетках, чему немало помогла и счастливо позаимствованная у гигантского вируса интеграза. Теперь, чтобы в результате отбора получился транспозон, защищающий клетку от вирусной инфекции, требовалось полностью или частично избавиться от промежуточного вируса и заставить предка Мавируса существовать самостоятельно. Мавирус, как это ни удивительно, может проникать в клетку Cafeteria roenbergensis и без посредничества CRoV. У него имеются для этого ферменты, а сам процесс был зафиксирован под микроскопом (электронным, разумеется). Так что в результате взаимовыгодных превращений получился ДНК-фрагмент, обладающий интегразой и способный самостоятельно проникать в эукариотическую клетку. Это и есть наш МП-транспозон.
Источник: Matthias G. Fischer, Curtis A. Suttle. A Virophage at the Origin of Large DNA Transposons // Science. 8 April 2011. V. 332. P. 231–234.
Елена Наймарк
![]()