![]()
Хромосомные инверсии ускоряют симпатрическое видообразование
|

Американские ученые экспериментально доказали, что хромосомные инверсии способствуют симпатрическому видообразованию. Они подавляют рекомбинацию набора генов с полезными мутациями, потому варианты с инверсией получают преимущество и быстро распространяются в подходящих местообитаниях. Материалом для доказательства послужили популяции губастика крапчатого, которые, получив инверсию, захватившую гены сроков цветения, разделились сообразно местным климатическим условиям.
Симпатрическое видообразование — это зарождение двух или нескольких видов на одной территории. Ему противопоставляют аллопатрическое видообразование, при котором для образования двух новых видов необходимо разделить их территориально с обязательных присутствием изолирующего барьера. Изолирующий барьер призван не давать скрещиваться разъединенным частям популяции, чтобы с двух сторон успели накопиться мутации. Они-то и определят в конечном итоге генетическую видоспецифичность и нескрещиваемость видов.
До конца прошлого века бытовало убеждение, что симпатрическое видообразование — это экзотика, редкое исключение на фоне ярких примеров аллопатрии. Однако чем дальше и внимательнее ученые исследуют видообразование, тем понятнее становится, насколько широко распространена симпатрия (см. новости на «Элементах»: Для видообразования не нужны барьеры, 13.02.2006; Эволюция без преград: ботаники нашли новое доказательство видообразования без географических барьеров, 13.02.2006, Видообразование на разных островах идет параллельными путями, 15.03.2007 и т. д.).
Начинает проясняться и механизм симпатрического видообразования. Это может быть неслучайное скрещивание носителей мутации — полезной или нейтральной (см.: Видообразование — личное дело каждого, «Элементы», 15.02.2006). Виды могут возникать также в результате неполной репродуктивной изоляции, которая не дает адаптивного преимущества никому из потомства, а потомство расходится просто в силу статистических законов или избирательной настройки половых партнеров на «свои» аллели (см.: Зафиксирован начальный этап видообразования у тропических бабочек, «Элементы», 09.11.2009). При этом пока что остаются малоизученными вопросы генетических механизмов симпатрического видообразования. Этот пробел будет постепенно заполняться, пример тому — недавняя публикация в журнале Plos Biology.
Дэвид Лоури и Джон Уиллис из Медицинского центра Университета Дьюка (Дарем, США) исследовали генетическое и экологическое разнообразие цветков с забавным названием «губастик крапчатый» (Mimulus guttatus). Это растение, родом из западных районов Северной Америки, сейчас распространено по всему североамериканскому континенту, завезено и в Старый Свет, и в Новую Зеландию, где успешно произрастает. В Северной Америке встречаются две формы этого вида губастика.
Одна из форм — однолетник, растущий в более или менее сухих местах. Цветет ранней весной. Вторая представлена многолетними растениями, которые многочисленны во влажных, тенистых прибрежных полосах. Время цветения сдвинуто на летний сезон, а ранняя весна предназначена для накопления зеленой массы. Эти два экотипа, очевидно, разобщены по времени цветения, и это препятствует их скрещиванию, то есть имеет место предзиготическая изоляция (см. репродуктивная изоляция).
Многолетник, пересаженный (искусственно или естественным образом распространившийся) в сухое местообитание, неизбежно проигрывает своим однолетним родичам. Его цветение приходится на самое сухое летнее время года, так что шансов оставить достаточное количество жизнеспособных семян у него почти нет. Однолетники во влажных местообитаниях, бросая все силы на раннее цветение, не успевают нарастить достаточно зеленой биомассы, так что позднецветущие родичи не оставляют им свободного места для роста. Кроме того, в прибрежных местообитаниях верхний слой почвы вместе с проростками и семенами смывается, так что удерживаются лишь те растения, которые уже имеют развитую корневую систему, а именно — многолетники.
Таким образом, нам ясно, что однолетняя весеннецветущая форма губастиков приспособлена как нельзя лучше к сухим местообитаниям, а многолетняя позднецветущая — к влажным прибрежным. Как шло становление этих различий, ученые выяснили в ходе генетического анализа, совмещенного с экспериментальной ботанической работой.
В распоряжении исследователей имелся обширный материал, характеризующий различные популяции как однолетней формы, так и многолетней (см. карту). Представители двух экотипов, как выяснилось, имеют инверсии. Одна из инверсий включает гены, связанные со временем цветения и некоторыми чертами морфологии.
|
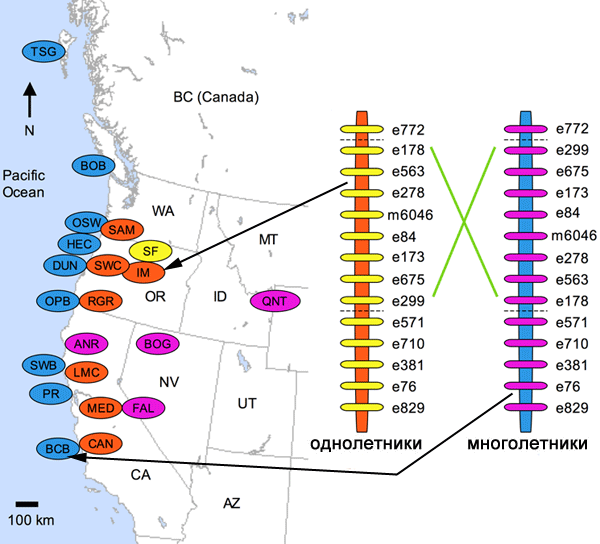
Насколько именно эта инверсия определила приспособление к разным местообитаниям? Чтобы ответить на этот вопрос, был предложен довольно трудоемкий, но исполнимый экспериментальный путь. Для этого серией гибридизации и обратного скрещивания были выведены особые формы, несущие все гены однолетников, но с инверсией многолетников. А также обратный вариант — с генами многолетников, но с инверсией однолетников. Далее была оценена приспособленность этих гибридов в условиях, оптимальных для однолетников (подсчитывалось количество и качество цветков, процент выживших растений и некоторые морфологические признаки). Растения с инверсией, как у многолетников, зацветают в среднем на 6 дней позже, каким бы ни был их генотип. Различие в приспособленности оказалось наивысшим для генотипа многолетников. У них «родная» инверсия снижает шансы на выживание в сухих условиях по сравнению со вставкой однолетников примерно в 9 раз. Так что для многолетнего экотипа жизненно важно иметь родную инверсию.
Ученые подчеркивают, что для симпатрического видообразования инверсии могут играть исключительно важную роль. Они подавляют рекомбинацию инвертированных участков генома. Если инверсия включает набор генов, которые полезны для выживания в конкретных условиях, то этот набор будет оставаться более или менее целостным. Он не будет рассеиваться по геному, смешиваясь с другими вредными или бесполезными аллелями. Таким образом, отбор будет преимущественно сохранять особей с этой исключительно удобной инверсией, так что она быстро распространится по популяции.
Для данного конкретного примера с губастиками очевидно, что инверсия захватила участок генома с несколькими генами, ответственными за время цветения и другие адаптивные признаки. Далее эта инверсия быстро распространилась среди тех популяций, где была уместна, вытеснив менее удачливые аллели. Параллельно менялись и другие гены в разошедшихся популяциях. В связи с этим ученые предполагают найти еще не одну полезную инверсию, ускорившую видообразование у губастика. По их мнению, инверсии являются мощным генетическим средством, ускоряющим видообразование.
Источники:
1) David B. Lowry, John H. Willis. A Widespread Chromosomal Inversion Polymorphism Contributes to a Major Life-History Transition, Local Adaptation, and Reproductive Isolation // PLoS Biol. 2010. 8(9): e1000500. Doi:10.1371/journal.pbio.1000500.
2) Mark Kirkpatrick. How and Why Chromosome Inversions Evolve // PLoS Biol. 2010. 8(9): e1000501. Doi:10.1371/journal.pbio.1000501.
Елена Наймарк
![]()