![]()
Избыточные регуляторы делают развитие эмбриона помехоустойчивым
|

Работа многих генов, управляющих развитием эмбриона, контролируется «теневыми» энхансерами — дополнительными регуляторными элементами, функция которых на первый взгляд кажется избыточной, потому что они дублируют действие основных регуляторных элементов («обычных» энхансеров). На примере гена svb, управляющего развитием щетинок у личинок дрозофилы, удалось показать, что теневые энхансеры являются «лишними» только в оптимальных условиях. При температурном стрессе и при мутациях «вышестоящих» регуляторных генов теневые энхансеры обеспечивают стабильность работы подконтрольного гена, тем самым придавая индивидуальному развитию помехоустойчивость.
Об открытии теневых энхансеров и об их эволюционной роли рассказано в заметке Эволюция регуляторных последовательностей ведет к эволюции видов («Элементы», 17.09.2008). Напомним, что энхансеры — это некодирующие регуляторные участки ДНК, располагающиеся, как правило, недалеко — в пределах нескольких десятков тысяч пар нуклеотидов (п.н.) — от начала контролируемого ими гена. Энхансеры содержат места прикрепления (сайты связывания) регуляторных белков — транскрипционных факторов. Прикрепляясь к энхансерам, транскрипционные факторы усиливают или ослабляют активность (экспрессию) соответствующего гена.
Теневые энхансеры характерны для многих генов, управляющих эмбриональным развитием. Они располагаются значительно дальше от начала контролируемого гена, чем «обычные» энхансеры, но функции выполняют те же самые. Теневой энхансер обычно «срабатывает» (то есть включает свой ген) в тех же клетках эмбриона и в то же время, что и основной энхансер. Такая избыточность генной регуляции, по-видимому, способствует эволюционным преобразованиям. В теневых энхансерах мутации накапливаются значительно быстрее, чем в основных (подробнее см. в вышеупомянутой заметке). Дублирование регуляторных участков ДНК дает возможность «избыточным» участкам более свободно накапливать мутации — точно так же, как это происходит в случае дупликации целых генов (см.: Многофункциональные гены — основа для эволюционных новшеств, «Элементы», 30.06.2008).
Впрочем, наличие теневых энхансеров у многих генов — регуляторов эмбрионального развития нельзя объяснить только тем, что они дают больше свободы для эволюционных преобразований. Естественный отбор не умеет заглядывать в столь отдаленное будущее. Теневые энхансеры должны выполнять какую-то полезную функцию здесь и сейчас, иначе они бы быстро разрушились под грузом мутаций и не встречались бы с таким постоянством около генов — регуляторов онтогенеза.
Выяснить эту функцию удалось американским и французским генетикам в ходе изучения гена svb (shavenbaby), который управляет формированием щетинок у личинки дрозофилы. Название гена, означающее в переводе «бритый младенец», показывает, что происходит, если ген выходит из строя. Регуляторная область гена svb интегрирует «входные сигналы» от нескольких генно-регуляторных каскадов. В результате совместного действия нескольких транскрипционных факторов, связывающихся с расположенными в этой области энхансерами, ген svb включается в тех клетках эмбрионального эпидермиса, которые должны дать начало щетинкам.
В некодирующей области длиной в 50 тыс. п.н. перед началом гена svb расположены три энхансера (их условные обозначения: 7, E и A). Их работа была изучена ранее (о методах поиска энхансеров и выявления их функций рассказано в заметке Генетические механизмы формирования сложных признаков постепенно проясняются, «Элементы», 14.04.2010; в обсуждаемой работе применялись схожие методы). Оказалось, что совместное действие энхансеров 7, E и A полностью объясняет распределение областей экспрессии гена svb в эпидермисе эмбриона: ген включается там, где срабатывает хотя бы один из трех энхансеров.
|
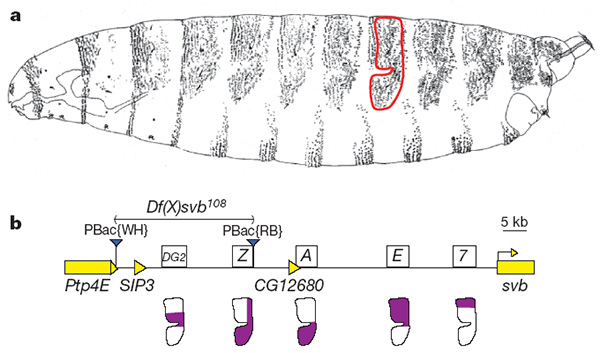
Казалось бы, ген svb не нуждается ни в каких дополнительных регуляторах, кроме энхансеров 7, E и A. Однако авторы обнаружили, что у него есть два теневых энхансера, расположенных на большем удалении от кодирующей части гена. Эти энхансеры получили название DG2 и Z. Функция новооткрытых энхансеров полностью перекрывается с функцией трех регуляторных элементов, известных ранее (см. рисунок).
Авторы продемонстрировали, что оба теневых энхансера эффективно работают (то есть включают присоединенный к ним ген там, где должны образоваться щетинки) у тех видов дрозофил, у которых есть щетинки этого типа (D. melanogaster, D. simulans) и не работают (испорчены мутациями) у вида, который в ходе эволюции утратил эти щетинки (D. sechellia). Три «обычных» энхансера у последнего вида тоже не работают.
Зачем нужны «избыточные» энхансеры? Чтобы ответить на этот вопрос, авторы удалили из генома мух участок ДНК, содержащий теневые энхансеры. На рисунке этот удаленный фрагмент обозначен двумя синими треугольничками; над ним написано «Df(X)svb108» — так назвали генно-модифицированную линию мух.
Удаление теневых энхансеров почти не повлияло на развитие щетинок: изменились только мелкие детали их строения. На здоровье личинок это не отразилось вовсе. Тем самым, казалось бы, подтвердилось предположение об «избыточности» теневых энхансеров. Однако дальнейшие эксперименты показали, что избыточность генной регуляции в данном случае имеет важное приспособительное значение: она повышает стабильность (помехоустойчивость) эмбрионального развития в неблагоприятных условиях.
Личинки с удаленными теневыми энхансерами, как выяснилось, развиваются нормально только при температуре, оптимальной для личинок дрозофил (около 25°C). Если же понизить температуру до 17 или повысить до 32°, то щетинки у мутантных личинок развиваются хуже. Авторы отмечают, что ген svb очень удобен для таких исследований, потому что число щетинок на том или ином участке эпидермиса в точности отражает уровень экспрессии этого гена. Нормальные личинки, из генома которых теневые энхансеры не были удалены, имеют одинаковое количество щетинок при любой температуре от 17 до 32°. Многочисленные дополнительные эксперименты подтвердили вывод о том, что теневые энхансеры стабилизируют онтогенез, обеспечивая достаточно высокий уровень экспрессии гена svb независимо от колебаний температуры.
Нормальный ход развития может быть нарушен не только внешними неблагоприятными воздействиями, такими как температурные колебания, но и внутренними — например, мутациями важных регуляторных генов. Могут ли теневые энхансеры «защищать» работающий ген и от таких помех? Чтобы проверить это, авторы использовали мух, гетерозиготных по мутации, выводящей из строя важнейший регуляторный ген wingless (см. об этом гене в заметке Генетические механизмы формирования сложных признаков постепенно проясняются, «Элементы», 14.04.2010). Ген svb находится под контролем нескольких регуляторных генов, в том числе wingless.
Оказалось, что для мух, у которых одна из двух копий wingless не работает, наличие теневых энхансеров является необходимым условием формирования нормальных щетинок. Если теневые энхансеры удалены, щетинки развиваются значительно хуже.
Таким образом, теневые энхансеры обеспечивают устойчивую работу гена svb не только при колебаниях температуры, но и при мутационном повреждении «вышестоящего» генно-регуляторного каскада.
Теневые энхансеры с «избыточными» функциями обнаружены в окрестностях многих генов — регуляторов развития не только у дрозофил, но и у мышей. Очень может быть, что это общее свойство данной группы генов у всех животных. Адаптивное (приспособительное) значение высокой помехоустойчивости онтогенеза очевидно. Поэтому отбор, как правило, должен способствовать закреплению мутаций, ведущих к росту независимости результатов развития от возмущающих воздействий. По-видимому, появление дополнительных энхансеров (например, в результате дупликации регуляторных участков ДНК) является одним из наиболее «легких» (вероятных) путей достижения этой цели.
Интересно, что побочным эффектом стабилизации онтогенеза может стать рост эволюционной пластичности. В нестабильных условиях отбор способствует формированию дополнительных энхансеров. Однако в стабильных благоприятных условиях эти энхансеры оказываются «лишними» и могут относительно свободно накапливать мутации. Если условия снова станут неблагоприятными, произошедшие в энхансерах изменения могут привести к дестабилизации онтогенеза и появлению каких-нибудь новых признаков (см.: Дестабилизация развития — путь к эволюционным новшествам, «Элементы», 13.07.2009). Известно, что изменения регуляторных последовательностей, управляющих работой генов — регуляторов развития, действительно играют ключевую роль в эволюции животных (см.: Прочтение генома опоссума доказало ключевую роль транспозонов в эволюции млекопитающих, «Элементы», 13.05.2007). Поэтому новые факты, обнаруженные авторами обсуждаемой статьи, могут оказаться очень важными для понимания эволюции животных.
Разумеется, теневые энхансеры — далеко не единственный механизм повышения помехоустойчивости онтогенеза. Этой же цели могут служить, например, контуры отрицательной обратной связи в генно-регуляторных сетях (Н. А. Колчанов, В. В. Суслов. Кодирование и эволюция сложности биологической организации), специализированные белки-шапероны (Белок Hsp90 контролирует активность мобильных генетических элементов, «Элементы», 19.01.2010) и микро-РНК (Li et al., 2009. A MicroRNA Imparts Robustness against Environmental Fluctuation during Development).
Источник: Nicolas Frankel, Gregory K. Davis, Diego Vargas, Shu Wang, Franзois Payre, David L. Stern. Phenotypic robustness conferred by apparently redundant transcriptional enhancers // Nature. Advance online publication 30 May 2010.
![]()