![]()
Эволюция цветного зрения у африканских цихлид шла двумя путями
|

В ходе стремительного видообразования у африканских цихлид сформировались разнообразные системы цветного зрения, ориентированные на световые волны разной длины. Эволюция цветного зрения шла двумя путями. Изменение активности генов светочувствительных белков-опсинов позволяло перенастраивать цветовое восприятие внутри границ воспринимаемого спектра, а сами эти границы менялись благодаря изменениям кодирующих участков генов.
Рыбы-цихлиды больших африканских озер — превосходный объект для изучения быстрого симпатрического видообразования. Озера имеют разный возраст, но все они относительно молоды. Каждое озеро изначально было заселено небольшим числом видов рыб, которые стали быстро дивергировать, причем эволюция в каждом озере шла в значительной мере независимо. Как обычно бывает в подобных случаях, освоение сходных ниш вело к независимому появлению сходных жизненных форм в разных озерах.
В природе разные виды африканских цихлид обычно не скрещиваются между собой (собственно, поэтому они и считаются разными видами, а не разновидностями). Репродуктивная изоляция обеспечивается в основном брачными предпочтениями. По-видимому, одним из главных факторов, обеспечивших быстрое видообразование у цихлид, был половой отбор (см.: Женская привередливость способствует видообразованию, «Элементы», 25.12.2009).
В природе для предотвращения гибридизации вполне достаточно брачных предпочтений, поэтому дополнительные изолирующие механизмы цихлидам не требуются, и они могут не возникать очень долго. Генетическая несовместимость развивается пассивно, как побочный результат накопления различий в изолированных генофондах, и на это требуется много времени — несколько миллионов лет, иногда даже более 10 миллионов. Это было показано путем сопоставления экспериментов по межвидовой гибридизации с оценками времени расхождения видов на основе сравнения их ДНК, с учетом геологической истории озер. В лабораторных условиях представители разных видов и даже родов африканских цихлид легко скрещиваются друг с другом и дают плодовитое гибридное потомство. Любопытно, что чем больше прошло времени с момента расхождения скрещиваемых видов, тем больше появляется у гибридного потомства новых признаков, отсутствующих у обоих родителей. Это может быть одним из способов появления новизны в эволюции (см.: Stelkens et al., 2009. Phenotypic novelty in experimental hybrids is predicted by the genetic distance between species of cichlid fish).
|

Важную роль при выборе партнера играет зрение: самцы каждого вида имеют свой особый наряд, а самки превосходно разбираются в нюансах их окраски. Недавно было показано, что зрение у разных видов цихлид может быть специально «настроено» на спектральные характеристики брачного наряда: например, если самцы у данного вида синие, то и глаза наиболее чувствительны именно к синим оттенкам (см.: Terai Y. et al. Divergent selection on opsins drives incipient speciation in Lake Victoria cichlids // PLoS Biology. 2006. № 4. P. e433. Doi:10.1371/journal.pbio.0040433). Всё это делает африканских цихлид хорошим объектом для изучения эволюции цветного зрения.
«Элементы» уже рассказывали об эволюции цветного зрения у позвоночных (см.: Обоняние и цветное зрение в эволюции млекопитающих развивались в противофазе, «Элементы», 18.06.2008). Доступный нашему зрению мир красок определяется разнообразием опсинов — светочувствительных белков, которые синтезируются в колбочках. Чтобы приобрести способность воспринимать новые оттенки, не нужно даже менять что-то в мозге — достаточно добавить в сетчатку новый опсин (см.: Обезьян вылечили от дальтонизма при помощи генной терапии, «Элементы», 18.09.2009; Мышиный мозг готов увидеть мир по-человечески, «Элементы», 29.03.2007). На примере опсинов удобно изучать эволюцию, поскольку в данном случае имеет место практически однозначное соответствие между генами и фенотипическими признаками (особенностями цветового восприятия).
Исследователи из США, Австралии и Швейцарии опубликовали в журнале PLoS Biology результаты широкомасштабного исследования опсинов у 54 видов цихлид из озера Малави и 11 видов из озера Виктория. У цихлид имеется один ген «черно-белого» опсина палочек (Rh1) и целых шесть классов «цветных» опсинов колбочек: SWS1 (ультрафиолетовый), SWS2B (фиолетовый), SWS2A (синий), Rh2B (голубой, или сине-зеленый), Rh2A (зеленый), LWS (красный). Наше унаследованное от обезьян трихроматическое зрение, вероятно, позволяет нам видеть лишь бледную тень того богатства красок, которое доступно цихлидам с их гексахроматическим зрением. У цихлид сетчатка имеет более сложное строение, чем у млекопитающих. Колбочки делятся на два морфологически различимых класса: «простые» и «двойные». В простых колбочках экспрессируются (работают) коротковолновые гены SWS, в двойных — длинноволновые Rh2 и LWS.
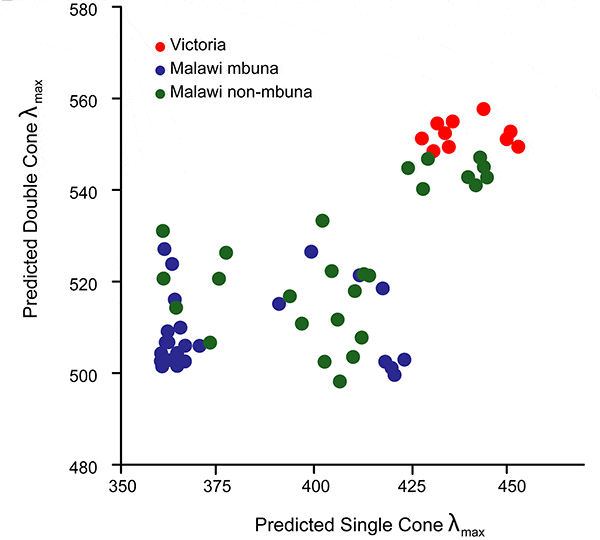
Авторы измерили активность опсиновых генов у всех изученных видов в обоих типах колбочек. Оказалось, что разные виды цихлид, в том числе и близкородственные, сильно различаются по относительной активности шести опсиновых генов. В озере Малави, где вода прозрачная, цихлиды используют все шесть генов, хотя и в очень разной степени. Малавийские виды распадаются на три группы, сетчатка которых наиболее чувствительна к коротким, средним и длинным световым волнам (рис. 3). В озере Виктория, где вода мутная, цихлиды почти не используют гены «ультрафиолетового» и «сине-зеленого» опсинов. У всех изученных цихлид из Виктории сетчатка наиболее чувствительна к длинноволновому свету, который лучше проходит сквозь мутную воду. Напомним, что «ультрафиолетовый» опсин является самым коротковолновым из опсинов простых колбочек, а «сине-зеленый» — самый коротковолновый из опсинов двойных колбочек.
|
Эти различия между озерами, по-видимому, отражаются и на окраске рыб: судя по рис. 1 и 2, у рыб из Малави преобладают синие и фиолетовые тона, у рыб из Виктории — красные и желтые.
Авторы сопоставили активность опсиновых генов с экологическими характеристиками изученных видов, а именно с питанием и типом местообитания. Оказалось, что только у одного гена («ультрафиолетового») активность тесно связана с типом питания. Среди рыб озера Малави наибольшая активность «ультрафиолетового» гена характерна для видов, питающихся фито- и зоопланктоном или водорослями, наименьшая — для хищников, питающихся другими рыбами или донными беспозвоночными. Эта особенность явно имеет адаптивный (приспособительный) характер, потому что, как было показано ранее, способность воспринимать свет в ультрафиолетовом диапазоне повышает эффективность питания у рыб–планктофагов. Все эти типы питания встречаются и у цихлид озера Виктория, однако «ультрафиолетовый» ген ни у кого из них не работает — ни у планктофагов, ни у хищников, потому что мутная вода Виктории непрозрачна для ультрафиолета.
В озере Виктория, как выяснилось, спектр активности опсиновых генов связан с мутностью воды в конкретных местообитаниях, а также с глубиной, на которой предпочитают жить представители данного вида. Чем мутнее вода и чем глубже обитают рыбы, тем слабее у них работает «фиолетовый» опсиновый ген, самый коротковолновый после «ультрафиолетового». В озере Малави четкой связи между локальной прозрачностью воды или глубиной обитания и экспрессией опсиновых генов обнаружить не удалось.
Авторы также изучили межвидовые различия в аминокислотных последовательностях опсинов, влияющие на их чувствительность к волнам разной длины. Оказалось, что эти различия распределены между шестью опсинами очень неравномерно. В озере Малави самым вариабельным оказался «ультрафиолетовый» опсин, на втором месте — «красный». В озере Виктория наибольшая вариабельность аминокислот, влияющих на избирательность цветового восприятия, отмечена у «красного» опсина. Выявленные различия меняют оптимум восприятия красного опсина на 5–15 нм. У видов, обитающих на больших глубинах, где доступная цветовая гамма сильно смещена в длинноволновую сторону, туда же смещена и чувствительность красного опсина. Аминокислотные различия, очевидно, закрепились не случайно, а под действием отбора (об этом свидетельствует, в частности, отсутствие корреляции между числом значимых и «молчащих» нуклеотидных замен в шести опсиновых генах).
Таким образом, эволюция систем цветового восприятия у африканских цихлид шла параллельно двумя путями: за счет изменения активности опсиновых генов и за счет изменения их кодирующих участков. Первым способом достигалась тонкая подстройка восприятия под особенности среды и образа жизни. Второй способ помогал менять границы воспринимаемого спектра — именно поэтому изменение аминокислотных последовательностей происходило преимущественно у двух «крайних» опсинов, ультрафиолетового и красного. В прозрачном озере Малави на эволюцию цветового зрения в большей степени влиял тип питания, в Виктории — спектральные характеристики доступного рыбам света, зависящие от мутности воды и глубины обитания.
Авторы подчеркивают, что столь высокое разнообразие систем цветового восприятия в пределах одной группы близкородственных родов и видов — случай уникальный, не имеющий аналогов у других позвоночных. Самое удивительное, что все эти контрастные различия развились очень быстро: озеру Малави 1–2 млн лет, озеру Виктория — не более 120 тысяч лет. Наблюдаемые различия в уровнях активности опсиновых генов у цихлид смещают оптимум цветового восприятия очень сильно — до 100 нм. Разные виды африканских цихлид действительно видят мир в разных красках. Это может влиять как на их поведение, так и на эволюцию других признаков — прежде всего их собственной окраски. Давно известно, что африканские цихлиды — самая разнообразная по окраске группа пресноводных рыб (не случайно их так любят аквариумисты). Теперь мы узнали, что и цветовое зрение у этих рыб тоже удивительно разнообразно.
Ранее было показано, что видообразование у цихлид в озере Виктория происходит под действием так называемого «сенсорного смещения» или «сенсорного драйва» (Seehausen et al. Speciation through sensory drive in cichlid fish // Nature. 2008. V. 455. P. 620–626). Так называют влияние особенностей органов чувств животных на направленность полового отбора. Например, если данный вид лучше всего видит красные объекты, самцам выгодно быть красными, чтобы привлекать внимание самок. Им выгодно также красоваться перед самками в таких местах (на такой глубине), где их краснота смотрится наиболее эффектно, более заметна для самок. Если интенсивность окраски у самца в какой-то мере отражает его здоровье и качество генов, то есть является «индикатором приспособленности» (а это обычно так и бывает), то самкам выгодно научиться еще лучше различать оттенки красного. Самки с наилучшей способностью различать эти оттенки выберут лучших самцов и оставят больше потомства, причем их сыновья унаследуют «удачный» брачный наряд от отца, а дочери — пристрастие к таким нарядам от матери.
В результате эволюция под действием полового отбора может приобрести черты автокаталитического процесса: рост интенсивности окраски будет стимулировать рост избирательности самок, и наоборот (см.: Fisherian runaway). Это может завести вид очень далеко по пути развития яркого наряда и изощренной системы цветового восприятия. Вряд ли можно сомневаться, что практически всё наблюдаемое у африканских цихлид буйство красок возникло под действием полового отбора, ведь для выживания вся эта пестрота не имеет существенного значения.
Источник: Christopher M. Hofmann, Kelly E. O'Quin, N. Justin Marshall, Thomas W. Cronin, Ole Seehausen, Karen L. Carleton. The Eyes Have It: Regulatory and Structural Changes Both Underlie Cichlid Visual Pigment Diversity // PLoS Biology. V. 7(12): e1000266. Doi:10.1371/journal.pbio.1000266.
![]()