![]()
Доказано участие гетерохроматина в видообразовании
|
Гетерохроматин — это участки нуклеотидных последовательностей, содержащие большей частью нейтральные повторы, а не уникальные белок-кодирующие гены. Эти участки обладают высокой изменчивостью. Однако до сих пор роль этих участков в видообразовании не была подтверждена. В серии изящных экспериментов американским ученым удалось доказать, что гетерохроматиновые участки хромосом у межвидовых гибридов ответственны за неправильное расхождение хромосом. Именно это вызывает летальность межвидовых гибридов и обеспечивает быстро возникающую репродуктивную изоляцию. Описанный механизм репродуктивной изоляции не требует наличия дополнительных изоляционных барьеров для видообразования.
ДНК, как всем хорошо известно, является основой наследственности и изменчивости. Разные виды обязаны иметь отличия в генетических последовательностях. Эти отличия появляются в результате мутаций, далее должны сработать механизмы, обеспечивающие концентрацию мутантных аллелей и сокращение смешиваемости носителей разной генетической информации. Сейчас известно множество способов, которые препятствуют скрещиванию особей с различной генетической информацией. Один из таких путей — это низкая жизнеспособность или летальность гибридов. Бывает и так, что нежизнеспособным оказывается только один пол у гибридов, а другой вполне нормально развивается. Ученые из Корнеллского университета (Итака, США) исследовали явление летальности гибридов именно с таким различием по полам. Свое исследование они провели на гибридах разных видов дрозофил. С одной стороны, дрозофила — это очень удобный объект, так можно исследовать генетическую подоплеку почти любого явления. С другой стороны, дрозофилы представляют необычный случай летальности гибридов. По правилу Холдейна, для гибридов нежизнеспособен тот пол, который несет две разные половые хромосомы (гетерогаметный пол, XY); а тот пол, который имеет одинаковые половые хромосомы (гомогаметный пол, XX) — выживает (см. также: Г. Э. Фельдман. «Джон Бэрдон Сандерсон Холдейн». Глава 2. «Научное творчество»). Но у дрозофил всё наоборот: у межвидовых гибридов выживают только гетерогаметные самцы, а гомогаметные самки умирают. Американские ученые разобрали механизм этого исключительного явления. Как выяснилось по ходу работы, во всём виноваты гетерохроматиновые участки хроматидных нитей.
|
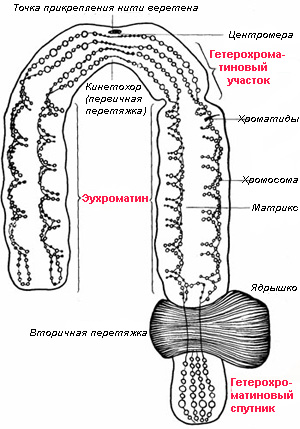
Гетерохроматин — это относительно плотно упакованные участки хроматидных нитей; этим он внешне отличается от эухроматина, имеющего рыхлую упаковку на протяжении большей части клеточного цикла.
Считается, что гетерохроматиновые участки не несут белок-кодирующих генов или таких генов очень мало; вместо них нуклеотидные последовательности гетерохроматина состоят большей частью из простых повторов. Гетерохроматин концентрируется у центромер, у теломер, образует сателлитные участки, а также периферический слой клеточного ядра. Гетерохроматин участвует в регуляции транскрипции генов, в особенности генов рибосомальных РНК, также играет важную роль в правильной постановке хромосом во время деления клетки. Несмотря на эти важные и, казалось бы, консервативные функции гетерохроматиновые участки чрезвычайно изменчивы и по обилию, и по составу даже у близких видов. Поэтому вполне логично заключить, что не только эухроматин, несущий белок-кодирующие гены, но и гетерохроматин как-то участвует и в видообразовании. И действительно, проведенные опыты доказали участие гетерохроматина в формировании стерильности гибридов.
Ученые скрестили самок Drosophila simulans и самцов Drosophila melanogaster. В результате скрещивания получились жизнеспособные оплодотворенные яйцеклетки. В них начиналось дробление, в течение первых трех часов после оплодотворения ядра делились 9 раз и формировалась обычная для плодовой мушки бластула с несколькими тысячами ядер без клеточных перегородок (так называемый многоядерный синцитий). По мере ядерных делений ядра нормально мигрировали к клеточной оболочке. В бластулах самцов нормальное деление продолжалось и дальше вплоть до 14-го деления, после которого бластодерма формировала клеточные мембраны, и синцитий становился нормальной многоклеточной бластулой. Вслед за этим начиналась гаструляция, и ученые получали жизнеспособных и плодовитых самцов.
Однако в бластулах самок после 9-го ядерного деления становились заметны нарушения. Ядра синцития располагались крайне неравномерно, оставляя пустыми большие участки клеточной периферии, но при этом скапливаясь комками в середине клетки. Ядра у клеточной периферии имели неправильную форму, неравномерно окрашивались, а это означало, что ядерные деления стали асинхронными. Кроме того, стали видны нарушения в расхождении хромосом в таких «неправильных» ядрах. Все бластулы, у которых наблюдались эти отклонения, оказались нежизнеспособными: их развитие останавливалось, многоядерная бластула не формировалась. Ясно, что именно этот дефект в нарушениях расхождения хромосом приводил к смерти эмбрионов.
|
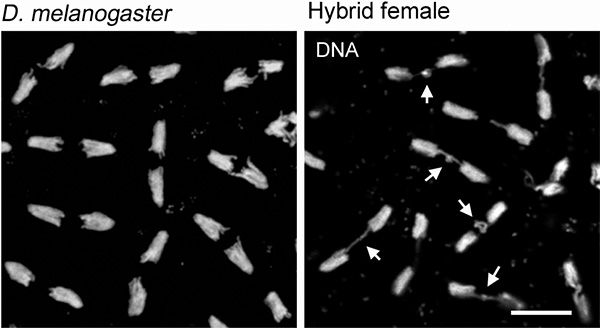
Что привело к подобным нарушениям в делении ядер и при чем тут гетерохроматин? Во-первых, ученым удалось выяснить, что ошибки получались только при расхождении X-хромосом Drosophila melanogaster. Иначе говоря, только X-хромосома, привнесенная самцом, являлась источником всех бед, а X-хромосома, полученная от самки Drosophila simulans, никак не влияла на процесс ядерного деления. Во-вторых, эти нарушения были связаны с нерасхождением особого гетерохроматинового участка X-хромосомы, который выявлялся с помощью специфического окрашивания.
Этот участок имеется у Drosophila melanogaster, но его нет у Drosophila simulans. Он содержит особый ген Zhr (zygotic hybrid rescue), который отвечает за гибель гибридных самок. Если этот ген отключить с помощью мутаций, то гибридные самки будут полностью жизнеспособны. У таких гибридов, как выяснилось, половые хромосомы расходятся совершенно нормально. Кроме того, на этом участке гетерохроматина находится специальный белок, который структурирует гетерохроматин между клеточными делениями, а во время клеточных делений равномерно распределяется по хроматину. У погибших гибридных самок этот белок так и оставался в скоплении на маленьком участке гетерохроматина на половой хромосоме. У эмбрионов Drosophila melanogaster simulans, как самцов, так и самок, и у гибридных самцов этот белок рассредотачивался во время клеточных делений.
Таким образом, круг поисков причин летальности гибридных самок сузился до небольшого участка гетерохроматина на половой хромосоме, а по времени — до 10–13-го ядерного деления. Именно во время этих делений и происходит формирование структуры гетерохроматина.
Итак если собрать воедино все эти факты, то причины, вызывающие смертность самок, проясняются. Общую модель процесса можно изобразить следующей схемой.
|
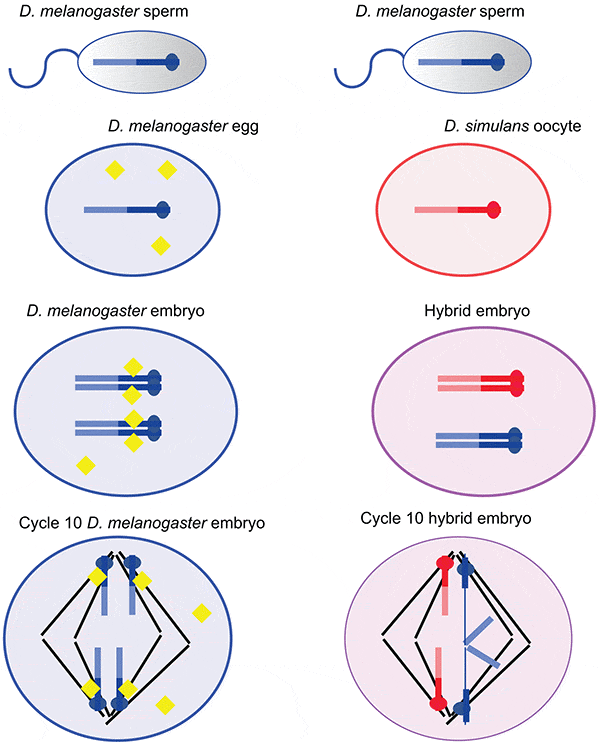
Участки нуклеотидных повторов в гетерохроматине могут быстро изменяться — они, как известно, чрезвычайно вариабельны. Если происходит такое изменение, то гибридные потомки рискуют остаться бесплодными или нежизнеспособными, и таким образом формируется репродуктивная изоляция. В этом случае для закрепления признака не потребуются специальные географические изоляционные барьеры, на которые сейчас принято делать основной упор. Репродуктивную изоляцию обеспечит измененная нуклеотидная последовательность, не несущая полезных или вредных мутаций в белок-кодирующих генах. Это случай репродуктивной изоляции, возникающей очень быстро при нейтральной мутации.
Источники:
1) Patrick M. Ferree, Daniel A. Barbash. Species-Specific Heterochromatin Prevents Mitotic Chromosome Segregation to Cause Hybrid Lethality in Drosophila // PLoS Biol 7(10): e1000234 (2009). doi:10.1371/journal.pbio.1000234.
2) Stacie E. Hughes, R. Scott Hawley. Heterochromatin: A Rapidly Evolving Species Barrier // PLoS Biol 7(10): e1000233 (2009). doi:10.1371/journal.pbio.1000233.
Елена Наймарк
![]()