![]()
Древо жизни завивается в кольца
|
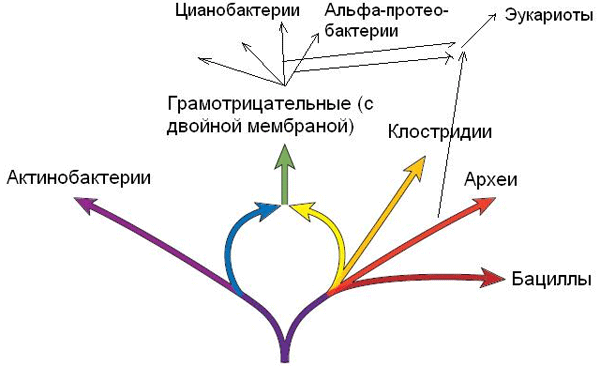
Известный специалист по эволюции микробов Джеймс Лейк из Калифорнийского университета привел веские аргументы в пользу того, что наиболее успешная и разнообразная группа прокариот, сыгравшая ключевую роль в развитии жизни на Земле, — грамотрицательные бактерии — возможно, возникла в результате слияния в единый организм двух более примитивных бактерий: актинобактерий и клостридий. Если эта гипотеза верна, схема ранней эволюции прокариот перестает быть похожей на дерево и приобретает вид кольца.
1. Дерево или клубок?
Со времен Дарвина эволюцию и родственные связи организмов схематически изображают в виде деревьев. «Древесная» форма эволюционных реконструкций основана на допущении об исключительно вертикальной природе наследственности (потомки получают все свои гены только от родителей). Однако в последние десятилетия стала ясна огромная роль горизонтальной передачи наследственного материала, когда организмы получают гены не только от родителей, но и от других организмов, в том числе и совершенно им не родственных (см.: Горизонтальный перенос генов и эволюция). Эволюционные реконструкции, построенные с учетом горизонтального генетического обмена, похожи не на деревья, а на спутанные клубки, сети или кольца (см.: На что похожа эволюция: на ветвящееся дерево или на сеть?).
Горизонтальный генетический обмен очень широко распространен у одноклеточных (особенно прокариот) и гораздо реже встречается у многоклеточных. Он может осуществляться множеством способов, включая перенос генов вирусами (см. трансдукция) и всасывание бактериями молекул ДНК из окружающей среды (см. трансформация). Но самым радикальным вариантом горизонтального генетического обмена является полное слияние геномов двух неродственных организмов.
2. Симбиогенез
Объединение двух геномов может произойти, в частности, в результате эндосимбиоза, то есть такого симбиотического объединения организмов, при котором один из них постоянно живет внутри другого.
Общепризнано, что именно таким — эндосимбиотическим — путем появились первые эукариоты (высшие организмы, в клетках которых имеется ядро). Геном эукариот исходно имеет химерную природу, поскольку он образовался путем слияния геномов нескольких неродственных прокариот, объединившихся в единый организм около 2 млрд лет назад (см.: А. В. Марков, А. М. Куликов. Происхождение эукариот как результат интеграционных процессов в микробном сообществе).
Теория симбиогенетического происхождения эукариот была неопровержимо доказана и стала общепризнанной лишь в последние десятилетия, а до этого научное сообщество долго отказывалось ее принимать. Достаточно сказать, что статья Линн Маргулис (Lynn Margulis) с первым убедительным изложением основ этой теории была отвергнута более чем дюжиной научных журналов, прежде чем ее опубликовал в 1966 году журнал, специализирующийся на спорных гипотезах (Journal of Theoretical Biology).
Судьба новой, не менее революционной гипотезы Джеймса Лейка, очевидно, будет далеко не столь драматичной, поскольку ее напечатал журнал Nature — благо Лейк ранее неоднократно публиковался в этом журнале и известен как крупный специалист по ранним этапам эволюции.
3. Гипотеза Лейка
По мнению Лейка, задолго до появления эукариот произошел еще один акт симбиогенеза, имевший весьма грандиозные последствия. Лейк утверждает, что крупнейшая и наиболее разнообразная группа прокариот — грамотрицательные бактерии — возникла в результате эндосимбиоза двух других, более древних и примитивных бактерий: актинобактерии (Actinobacteria) и клостридии (Clostridia).
Чтобы понять суть гипотезы, равно как и ее красоту и «революционность», нужно вкратце познакомиться с главными героями — грамотрицательными бактериями (они же — бактерии с двойной мембраной).
В настоящее время не существует общепринятой классификации прокариот. Однако, по мнению Лейка, прокариоты отчетливо подразделяются на пять больших естественных (то есть происходящих каждая от своего предка) группировок. Это археи, бациллы (и их родня), клостридии (и примкнувшие к ним), актинобактерии и грамотрицательные бактерии. Лейк не разделяет общепринятого мнения о том, что всех прокариот нужно делить сначала на архей и бактерий, а потом уже бактерий делить на более мелкие группировки. Результаты исследований Лейка и его коллег, основанные, в частности, на анализе делеций (выпавших кусков) и вставок в некоторых медленно эволюционирующих генах, свидетельствуют о том, что потомки «последнего общего предка всего живого» (см.: LUCA) разделились сначала не на предков бактерий и предков архей (как считает большинство), а на предков актинобактерий и на предков эволюционной линии, включающей клостридий, бацилл и архей (см. рис. 1).
Грамотрицательные бактерии занимают среди прокариот особое место, причем по ряду признаков их можно назвать «высшими», «наиболее эволюционно продвинутыми» или «самыми высокоорганизованными» прокариотами. Грамотрицательные бактерии очень многочисленны и разнообразны. К ним относятся цианобактерии, обогатившие земную атмосферу свободным кислородом около 2,4 млрд лет назад и давшие начало пластидам эукариот, а также разнообразные протеобактерии, в том числе альфа-протеобактерии, которые, по-видимому, первыми «изобрели» наиболее эффективный вариант кислородного дыхания и дали начало митохондриям эукариот. Главным отличием грамотрицательных бактерий от всех остальных (грамположительных) прокариот является более сложное строение клеточной оболочки. У грамположительных прокариот оболочка клетки состоит из одной липидной мембраны, снаружи от которой находится толстый слой пептидогликана. У грамотрицательных бактерий снаружи от пептидогликанового слоя находится еще одна мембрана, имеющая довольно сложное и специфическое строение. В учебниках по микробиологии обычно пишут, что внутренняя мембрана грамотрицательных бактерий гомологична единственной мембране грамположительных бактерий (то есть происходит от нее), а наружная возникла позже, как некое вторичное дополнение к исходно одномембранной клеточной оболочке более древних грамположительных бактерий.
С другой стороны, двойная мембрана неизбежно наводит на мысли об эндосимбиозе. Что если некая бактерия когда-то приобрела внутриклеточного симбионта — другую бактерию, и они впоследствии слились в единый организм? В этом случае можно ожидать, что получившаяся химерная клетка будет иметь двойную мембрану. Мембрана эндосимбионта станет внутренней мембраной объединенного организма, а мембрана хозяина — наружной. Но это, конечно, не доказательство, а лишь некий намек на возможное направление поиска. Основой для гипотезы Лейка стали совсем другие факты, полученные в ходе анализа распределения семейств белков по пяти основным группировкам прокариот.
4. Статистические обоснования гипотезы
Хотя практически все эксперты уже признали важнейшую роль горизонтального генетического обмена в эволюции прокариот, необходимые методологические и практические выводы из этого пока не сделаны. Это проявляется, прежде всего, в том, что весь мощнейший математический аппарат и сложнейшее программное обеспечение, разработанное специально для построения эволюционных реконструкций на основе молекулярно-генетических данных, до сих пор ориентированы на построение эволюционных деревьев, а вовсе не клубков, сетей или колец. Подавляющее большинство компьютерных программ, используемых биологами для воссоздания путей эволюции, получают на выходе нуклеотидные последовательности ДНК сравниваемых организмов (или другие их признаки), а на выходе выдают деревья — и ничего кроме деревьев. Получается, что на словах ученые уже признали, что эволюция прокариот не была древовидной, а на деле по-прежнему продолжают исходить из предположения о ее древовидности. Тем самым неявно допускается, что роль горизонтального переноса в эволюции прокариот была пренебрежимо мала — хотя неправомерность такого допущения уже очевидна для всех.
Джеймс Лейк одним из первых стал разрабатывать новые статистические методы, позволяющие делать осмысленный выбор между древовидными и «кольцевыми» эволюционными реконструкциями. Эти методы ранее были успешно проверены на эукариотах. В результате химерная природа эукариотического генома получила новое убедительное подтверждение, а схему, иллюстрирующую происхождение эукариот путем слияния нескольких разных прокариот в единый организм, стали называть «кольцом жизни» (в противовес общепринятому термину «древо жизни»; см.: Rivera, M. C. & Lake, J. A. The ring of life: evidence for a genome fusion origin of eukaryotes // Nature. 2004. V. 431. P. 152–155).
Теперь Лейк применил эти методы к пяти вышеперечисленным группам прокариот. Это позволило количественно оценить вероятность (правдоподобие) альтернативных эволюционных схем. Оказалось, что наилучшее из возможных деревьев имеет значительно более слабую статистическую поддержку, чем наилучшая из возможных схем с кольцами (см. рис. 2).
|
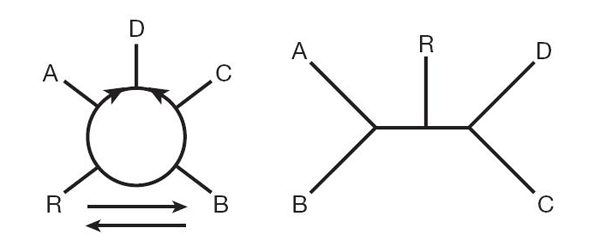
Автор опирался в основном на данные по наличию (или отсутствию) различных семейств белков в пяти группах прокариот. Наиболее информативными в данном случае являются такие семейства белков, которые есть у трех из пяти групп и отсутствуют у двух остальных. Всего существует 10 возможных вариантов распределения таких белковых семейств по пяти группам (есть у групп 1, 2, 3 и нет у групп 4 и 5; есть у групп 1, 2, 4 и нет у групп 3 и 5, и т. д.). Если верна левая (кольцевая) схема на рис. 2, то следует ожидать, что из этих 10 вариантов пять будут встречаться редко, а пять — часто. Часто должны встречаться такие семейства белков, которые есть одновременно у актинобактерий (A) и грамотрицательных (D) и (или) одновременно у клостридий (C) и грамотрицательных (D). Семейств белков, распределенных иначе, должно быть мало. Оказалось, что это предсказание кольцевой схемы выполняется с абсолютной точностью: все пять распределений, которые должны встречаться часто, действительно встречаются часто, и наоборот — все пять распределений, которые должны встречаться редко, действительно встречаются редко.
Древовидная схема (правая на рис. 2) дает несколько иные предсказания относительно частоты встречаемости каждого из 10 распределений, и эти предсказания существенно расходятся с реальностью.
На основе этого и ряда других статистических тестов Лейк делает вывод, что кольцевая схема, в которой грамотрицательные бактерии происходят путем слияния актинобактерии с клостридией, является намного более правдоподобной, чем любая из возможных древовидных схем.
5. Почему именно эндосимбиоз?
Статистические тесты, о которых говорилось в предыдущем разделе, фактически говорят лишь о том, что геном грамотрицательных бактерий имеет химерное происхождение, то есть образовался в результате объединения генов клостридий и актинобактерий. Это могло произойти не только путем эндосимбиоза, но и иными, менее драматическими путями. Например, какая-нибудь клостридия могла понемногу заимствовать гены актинобактерий до тех пор, пока не набрала их очень много. При этом строение клетки постепенно менялось, в том числе появилась и вторая мембрана. Разумеется, могло быть и наоброт: актинобактерия могла позаимствовать много генов у клостридий. Можно предположить и простое слияние клеток с последующим объединением геномов (хотя у современных прокариот, в отличие от эукариот, клетки сливаться не могут). Почему же Лейк считает, что имел место именно эндосимбиоз — то есть либо актинобактерия поселилась внутри клостридии, либо наоборот? Собственно, единственным весомым аргументом в пользу этого предположения является двойная мембрана грамотрицательных бактерий, о чем говорилось выше. Эндосимбиоз дает очень простое и красивое объяснение появлению двойной мембраны (наружная мембрана принадлежит хозяину, внутренняя — симбионту). У этой гипотезы есть проверяемые следствия — например, следует ожидать, что одна из двух мембран сохранила в себе что-то, характерное для клостридий, а вторая — для актинобактерий. Так что будущее обязательно покажет, верна ли догадка Лейка.
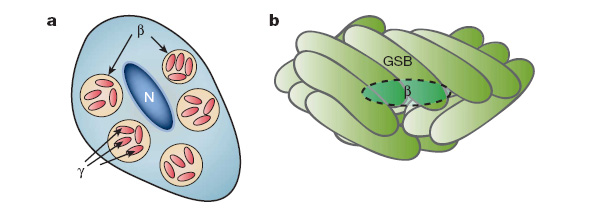
Слабым местом гипотезы является тот факт, что для современных прокариот эндосимбиозы не характерны. Этим прокариоты радикально отличаются от эукариот, которые многократно в ходе своей истории обзаводились разнообразными внутриклеточными симбионтами. Однако и прокариоты все-таки не совсем «безнадежны» в этом отношении. Например, есть веские основания полагать, что один из многочисленных внутриклеточных симбионтов тлей представляет собой двух «вложенных» друг в друга бактерий: бета-протеобактерию, внутри которой живут гамма-протеобактериальные симбионты (Carol D. von Dohlen et al. Mealybug ?-proteobacterial endosymbionts contain ?-proteobacterial symbionts // Nature. 2001. V. 412. P. 433–436). Чисто прокариотические эндосимбиозы неизвестны, однако описана одна чрезвычайно интересная симбиотическая система, которая показывает, с чего может начинаться такой симбиоз. Речь идет о бактериальном комплексе, известном под названием Chlorochromatium aggregatum. Центральный компонент этого комплекса представляет собой подвижную гетеротрофную (питающуюся органикой) бета-протеобактерию. Вокруг нее аккуратными стопками располагаются от 10 до 60 фотосинтезирующих зеленых серных бактерий. Все компоненты этого симбиотического комплекса соединены выростами наружной мембраны центральной бета-протеобактерии. Предполагается, что у всех компонентов комплекса имеется общее периплазматическое (межмембранное) пространство. Смысл этого содружества в том, что подвижная бета-протеобактерия перетаскивает всю компанию в места, благоприятные для жизни привередливых серных бактерий, а серные бактерии занимаются фотосинтезом и обеспечивают пищей и себя, и бета-протеобактерию. Заметим, что в обоих приведенных примерах все компоненты симбиотических комплексов являются грамотрицательными бактериями.
|
Если гипотеза Лейка верна, то целенаправленный поиск должен привести к тому, что новые доказательства посыплются, как из рога изобилия — как это происходит с симбиогенетической теорией происхождения эукариот со времен публикации статьи Маргулис. Сам Лейк вскользь указывает на одно дополнительное подтверждение, связанное с эволюцией фотосинтеза. Дело в том, что среди прокариот фотосинтез встречается только у некоторых клостридий, а также у многих грамотрицательных бактерий (цианобактерий, пурпурных протеобактерий, зеленых серных бактерий и др). Ни одна из древовидных эволюционных реконструкций не могла объяснить такое странное распределение способности к фотосинтезу среди прокариот — даже с учетом возможного горизонтального переноса. Для фотосинтеза необходима уйма генов, и приобрести их все разом путем обычного горизонтального переноса весьма трудно. Кольцевая схема Лейка, напротив, хорошо объясняет эту ситуацию. По-видимому, изобретателями фотосинтеза были какие-то древние клостридии. От этих клостридий фотосинтез достался по наследству первым грамотрицательным бактериям (которые, следовательно, изначально были фотосинтезирующими организмами). В дальнейшем во многих группах грамотрицательных бактерий фотосинтез был вторично утрачен. Отсюда — еще одно проверяемое следствие: можно ожидать, что, порывшись хорошенько в геномах нефотосинтезирующих грамотрицательных бактерий, мы найдем там следы древних фотосинтетических систем.
Гипотеза Лейка кажется очень логичной, она делает общую картину ранней эволюции более стройной и красивой. Получается, что важнейшее в истории земной жизни эволюционное событие — появление эукариот путем симбиогенеза — было подготовлено другим, более древним симбиогенетическим событием, которое к тому же создало предпосылки для обогащения атмосферы кислородом. Однако следует признать, что фактов, подтверждающих эту гипотезу, пока еще маловато. Будем надеятся, что публикация в Nature привлечет внимание исследователей к проблеме происхождения грамотрицательных бактерий, и скоро станет ясно, правильно ли угадал Лейк или попал пальцем в небо.
Источник: James A. Lake. Evidence for an early prokaryotic endosymbiosis // Nature. 2009. V. 460. P. 967–971.
См. также:
- А. В. Марков. Горизонтальный перенос генов и эволюция.
- Горизонтальный перенос генов приводит к новому режиму эволюции, «Элементы», 05.02.2007.
- На что похожа эволюция: на ветвящееся дерево или на сеть?
- А. В. Марков, А. М. Куликов. Происхождение эукариот как результат интеграционных процессов в микробном сообществе.
- Происхождение эукариот.
Александр Марков
![]()