![]()
Удвоение генов может приводить к видообразованию
|

Французские и британские генетики обнаружили простой механизм, при помощи которого между организмами, относящимися к одному виду, может быстро возникнуть частичная репродуктивная изоляция (нескрещиваемость), что, в свою очередь, в дальнейшем может привести к разделению исходного вида на два. Для возникновения изоляции достаточно, чтобы жизненно важный ген удвоился, а затем у разных представителей вида вышли из строя разные его копии.
Возникновение репродуктивной изоляции — ключевой этап процесса видообразования, и поэтому едва ли стоит удивляться тому огромному вниманию, которое биологи уделяют изучению изолирующих механизмов (см. ссылки внизу). Изоляция бывает презиготическая (организмы не хотят или не могут скрещиваться, или скрещиваются, но оплодотворения не происходит) и постзиготическая (оплодотворение происходит, но гибридное потомство погибает, оказывается бесплодным или имеет пониженную приспособленность).
Биологи давно предполагали, что одной из причин возникновения постзиготической изоляции может быть несовместимость аллельных вариантов нескольких взаимодействующих между собой генов. Теоретически между разными генами одного и того же генома может существовать такое взаимодействие (см.: epistasis), что жизнеспособными будут не все сочетания аллелей этих генов, а только некоторые. Например, генотипы A1A1B1B1 и A2A2B2B2 могут быть жизнеспособны, а A1A1B2B2 и A2A2B1B1 — нет. В этом случае носителям первых двух генотипов будет выгодно скрещиваться только с себе подобными, и отбор будет способствовать тому, чтобы они научились выбирать «правильных» партнеров. В конечном счете это может привести к формированию презиготической изоляции и разделению исходного вида на два. Однако до сих пор лишь очень небольшое число таких случаев удалось «расшифровать» на молекулярном уровне, то есть выявить конкретные пары взаимодействующих генов, у которых есть нежизнеспособные комбинации аллелей, и установить их функцию, нуклеотидную последовательность и природу взаимодействия.
Французские и британские генетики расшифровали один такой случай у классического модельного растения резушки Таля (Arabidopsis thaliana), которая лучше известна молекулярным биологам под своим латинским названием «арабидопсис».
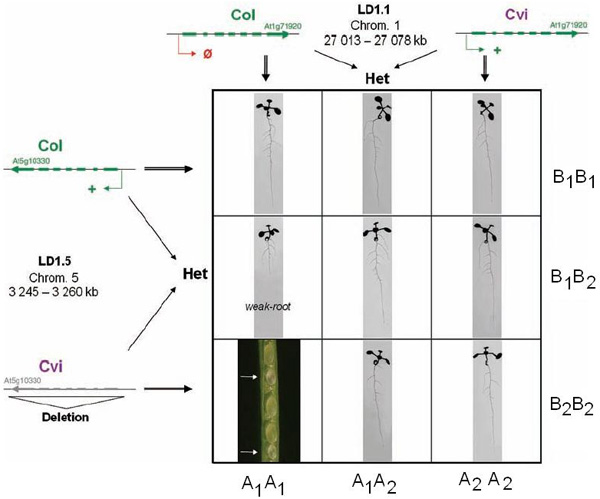
Ранее было замечено, что при скрещивании двух лабораторных линий арабидопсиса, происходящих от разных природных популяций (условные обозначения линий — Col и Cvi), в первом поколении гибридов никаких отклонений от нормы не наблюдается, но при дальнейшем скрещивании гибридов между собой часть образующихся семян оказывается нежизнеспособной (в семени не развивается зародыш). При помощи стандартных методов генетического анализа было установлено, что гибель семян объясняется существованием несовместимых комбинаций аллелей двух локусов (локус — участок хромосомы, где может быть расположен один или несколько генов). Один из локусов располагается на 1-й хромосоме (его назвали LD1.1), другой — на пятой (он получил название LD1.5).
Сначала исследователи стандартными методами генетического картирования (см.: генетические карты сцепления) сузили круг поиска, сократив размеры обоих «подозреваемых» участков ДНК до нескольких десятков тысяч пар нуклеотидов. Теперь можно было посмотреть, какие гены попали в эти участки. Оказалось, что в геноме линии Col в обоих участках находится по одной копии жизненно важного гена HPA. Этот ген кодирует фермент гистидинол-фосфат-амино-трансферазу, необходимый для синтеза аминокислоты гистидина. В статье используется весьма сложная номенклатура генов, мы же для простоты обозначим копию HPA, находящуюся в первой хромосоме, буквой A, а в пятой — буквой B. Аллельные варианты, характерные для линии Col, будем обозначать единицей, для линии Cvi — двойкой.
Вначале были исследованы аллели, характерные для линии Col, то есть A1 и B1. Их кодирующие последовательности отличаются друг от друга только двумя синонимичными нуклеотидными заменами, то есть белки, ими кодируемые, совершенно одинаковы. Наличие двух почти одинаковых копий одного и того же гена, судя по всему, является результатом недавней генной дупликации.
Затем исследователи определили уровень экспрессии (активность работы) генов A1 и B1. Для этого они выделяли из клеток матричные РНК (мРНК). Поскольку гены различаются двумя нуклеотидными заменами, считанные с них мРНК можно различить, хотя эти замены и не влияют на структуру белка, синтезируемого на основе мРНК. Оказалось, что ген A1 не работает (не считывается) ни в одной из тканей растения, тогда как ген B1 нормально работает и в корнях, и в побегах. Отсутствие активности у гена A1 объясняется, вероятно, какими-то пока еще не выявленными мутациями в регуляторных областях гена.
В линии Cvi ситуация оказалась обратной. У этих растений копия гена, расположенная в первой хромосоме (в нашей номенклатуре — A2), работает нормально, причем уровень активности у аллеля A2 немного выше, чем у B1. Что касается аллеля B2, то он оказался нефункциональным. Более того, в соответствующем участке пятой хромосомы в линии Cvi вообще не оказалось кодирующей последовательности гена HPA. Зато там обнаружились фрагменты мобильных генетических элементов. Это позволяет предположить, что копия гена HPA пропала из этого локуса в результате деятельности транспозонов. Можно, конечно, допустить, что ее там никогда и не было (то есть дупликация исходного гена произошла у предков линии Col, но ее не было у предков линии Cvi). Однако это не подтверждается результатами анализа других линий арабидопсиса, у которых практически всегда в соответствующем локусе пятой хромосомы есть копия гена HPA — иногда работающая, иногда нет.
Дальнейшие исследования подтвердили, что несовместимость при скрещивании двух линий арабидопсиса объясняется тем, что семена, в геноме которых собираются вместе нефункциональные аллели (генотип A1A1B2B2), оказываются нежизнеспособными. Остальные восемь возможных генотипов — жизнеспособны (см. рисунок). Иными словами, семя погибает, если в его геноме не функционирует ни одна из четырех копий гена HPA, что приводит к резкому недостатку гистидина в развивающемся зародыше. Правда, у растений с генотипом A1A1B1B2, имеющих только одну рабочую копию гена — B1, наблюдается недоразвитие корня. Это объясняется тем, что ген B1 экспрессируется слабее, чем A2, а корень — часть растения, наиболее чувствительная к недостатку гистидина. Если поливать растение с таким генотипом раствором гистидина, то есть вносить дефицитную аминокислоту извне, то корень вырастает нормальный. Более того, если подкармливать гистидином растения, у которых часть семян имеет «нежизнеспособный» генотип A1A1B2B2, то во всех семенах развивается нормальный зародыш.
Установив эти факты, исследователи проанализировали еще 30 линий арабидопсиса. Во всех линиях обнаружились две копии гена HPA, располагающиеся в одних и тех же локусах на первой и пятой хромосомах. При этом в большинстве случаев (22 из 30) одна из двух копий оказалась тем или иным способом выведена из строя. Авторы обнаружили шесть разных вариантов мутаций, выводящих из строя этот ген. Это либо отсутствие экспрессии, как в линии Col, либо делеция (выпадение) той или иной части кодирующей последовательности (в линии Cvi, как мы помним, выпала вся кодирующая последовательность), либо преждевременный стоп-кодон. Более четверти из всех возможных вариантов скрещиваний между линиями приводят к образованию нежизнеспособных семян.
По-видимому, произошедшее относительно недавно удвоение гена HPA привело к тому, что в пределах вида Arabidopsis thaliana наметилось разделение на две взаимно несовместимые группы популяций: те, у которых вышла из строя копия гена HPA на первой хромосоме, и те, у кого «поломалась» копия на пятой хромосоме. Очевидно, что при определенных условиях эти две группы популяций могут в будущем превратиться в отдельные виды.
|
Таким образом, стал ясен один из возможных сценариев относительно быстрого зарождения репродуктивной изоляции внутри вида. Сначала происходит удвоение какого-нибудь жизненно важного гена. Когда ген удваивается, естественный отбор перестает выбраковывать мутации, снижающие активность одной из его копий или выводящие ее из строя (потому что в запасе есть вторая копия, которая и в одиночку вполне справляется со своей функцией). В результате с большой вероятностью одна из двух копий гена становится нефункциональной.
Однако у разных представителей одного и того же вида могут выйти из строя разные копии удвоенного гена. При этом неизбежно возникает частичная несовместимость (постзиготическая репродуктивная изоляция) между популяциями, в которых распространились мутации, выводящие из строя разные копии гена. Поскольку скрещивания между представителями таких популяций становятся невыгодными (дают меньше потомков в первом или последующих поколениях), отбор будет благоприятствовать выработке любых механизмов, предотвращающих скрещивания с «неподходящими» партнерами. В случае животных это могут быть изменения в алгоритмах выбора брачного партнера; у растений аналогичный результат может быть достигнут, например, за счет изменения сроков цветения, взаимоотношений с насекомыми-опылителями или путем модификации биохимических механизмов «выбора пыльцы», осуществляемого тканями пестика.
Данный способ видообразования предполагался теоретиками уже давно, но так наглядно продемонстрировать его начальные этапы, вскрыв при этом молекулярные механизмы явления, удалось, пожалуй, впервые.
Источник: David Bikard, Dhaval Patel, Claire Le Mette, Veronica Giorgi, Christine Camilleri, Malcolm J. Bennett, Olivier Loudet. Divergent Evolution of Duplicate Genes Leads to Genetic Incompatibilities Within A. thaliana // Science. 2009. V. 323. P. 623–626.
О механизмах видообразования см. также:
1) Для видообразования достаточно одного гена, «Элементы», 28.11.2007.
2) Видообразование — личное дело каждого, «Элементы», 15.02.2006.
3) Расшифрован генетический механизм, позволяющий бактериям отличать «своих» от «чужих», «Элементы», 14.07.2008.
4) Начальные этапы видообразования воспроизведены в эксперименте на дрожжах, «Элементы», 06.06.2007.
5) Эволюция без преград: ботаники нашли новое доказательство видообразования без географических барьеров, «Элементы», 13.02.2006.
6) Для видообразования не нужны барьеры, «Элементы», 13.02.2006.
Александр Марков
![]()