![]()
Конечности у членистоногих и кольчатых червей растут из общего корня
|

Традиционно считалось, что членистоногие произошли от кольчатых червей, причем ноги первых развились из параподий (парных конечностей) последних. Сравнительный анализ ДНК опроверг мнение о близком родстве членистоногих и кольчецов, что заставило предположить независимое происхождение конечностей у этих двух типов животных. Однако новые эмбриологические данные показали, что конечности у них все-таки имеют общее происхождение. Из этого следует, что общий предок всех первичноротых был сегментированным животным с парными конечностями. Впоследствии конечности были утрачены во многих эволюционных линиях (в том числе у моллюсков и круглых червей).
«Элементы» уже рассказывали о радикальных изменениях в системе животного царства, которые произошли в связи с развитием методов молекулярной филогении — эволюционных реконструкций на основе сравнительного анализа ДНК (см.: Новые данные позволили уточнить родословную животного царства, «Элементы», 10.04.2008). Наиболее серьезные изменения произошли в систематике первичноротых (Protostomia), к которым по традиционным представлениям относились кольчатые черви, моллюски и членистоногие, а по новым данным — еще и круглые черви, плоские черви и ряд других групп.
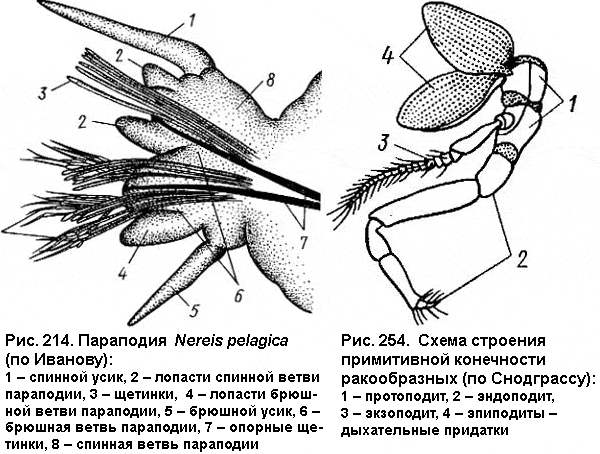
Самый большой казус вышел с членистоногими и кольчатыми червями. Эти два типа животных анатомически очень похожи друг на друга. Для них характерно членистое (сегментированное) тело с парой конечностей на каждом сегменте, причем строение конечностей во многом сходно (см. рисунок). В обоих случаях конечность исходно двуветвистая (состоит из спинной и брюшной ветви, или экзоподита и эндоподита), а сверху на ней часто имеются дыхательные (жаберные) придатки. Неудивительно, что специалисты по сравнительной анатомии уверенно выводили членистоногих из кольчецов, а конечности их считали гомологичными (имеющими общее происхождение): экзоподит конечности членистоногих выводился из спинной ветви параподии кольчецов и т. д.
Новые молекулярные данные заставили отказаться от этой стройной теории. Выяснилось, что кольчецы — родственники моллюскам (что признавалось и «классической» школой), но не членистоногим. Ближайшей родней членистоногих оказались круглые черви, чего никто не ожидал. Согласно новым представлениям, первичноротые очень рано разделились на две ветви: Ecdysozoa (членистоногие, круглые черви, тихоходки, онихофоры и др.) и Lophotrochozoa (кольчатые черви, моллюски, брахиоподы, немертины, форониды и по крайней мере часть плоских червей). Нашлись и немолекулярные признаки, подтверждающие новую классификацию: у Lophotrochozoa есть особая личинка-трохофора, у Ecdysozoa ее нет (ранее предполагали, что у предков членистоногих трохофора была, но потом «потерялась»); Ecdysozoa в процессе роста линяют, Lophotrochozoa растут без линек.—
Как же в этом случае объяснить явное сходство строения кольчатых червей и членистоногих, затрагивающее три важнейших признака (сегментация тела, парные конечности на сегментах, вторичная полость тела — целом)? По каждому из этих признаков возможны два варианта: либо кольчецы и членистоногие приобрели данный признак независимо друг от друга, либо признак имелся у общего предка первичноротых, сохранился у кольчецов и членистоногих, но был утрачен другими представителями Ecdysozoa и Lophotrochozoa. В первом случае названные структуры (сегменты, конечности, целом) у кольчецов и членистоногих являются гомологичными, во втором они негомологичны, то есть имеют разное происхождение.
|
Для решения вопроса о гомологичности органов или частей тела традиционно используются сравнительно-анатомические и эмбриологические данные. Например, если органы развиваются схожим образом из одинаковых зачатков, расположенных в одном и том же месте зародыша, то они, скорее всего, гомологичны. В последнее время большую помощь анатомам и эмбриологам оказывают данные по экспрессии генов — регуляторов развития (см.: Новое в науке о знаменитых Hox-генах, регуляторах развития, «Элементы», 10.10.2006). Если у двух зародышей в какой-то части тела экспрессируются одни и те же регуляторные гены — это важный довод в пользу гомологичности этих частей (и тех органов, которые из них затем разовьются).
Именно таким способом недавно удалось разобраться с проблемой сегментации тела у членистоногих и кольчецов (Prud'homme et al., 2003. Arthropod-like Expression Patterns of engrailed and wingless in the Annelid Platynereis dumerilii Suggest a Role in Segment Formation; Saudemont et al., 2008. Complementary striped expression patterns of NK homeobox genes during segment formation in the annelid Platynereis). Оказалось, что в обоих типах животных закладка сегментов идет под управлением одних и тех же генов, из которых одни экспрессируются в передней части будущего сегмента, а другие — в задней. В результате закладываются первичные сегменты, которые предложено называть «парасегментами». У кольчецов первичная сегментация сохраняется на всю жизнь, иными словами, сегменты взрослого червя соответствуют парасегментам зародыша. У членистоногих происходит перекройка первичных сегментов: границы парасегментов сглаживаются, а затем возникают границы «взрослых» сегментов, не соответствующие старым границам (передняя часть каждого парасегмента присоединяется к парасегменту, расположенному спереди). Таким образом, парасегменты зародышей кольчецов и членистоногих гомологичны друг другу, а сегменты взрослых животных — нет. Каждый сегмент тела членистоногих образуется из частей двух парасегментов.
Из этого был сделан вполне логичный вывод, что общий предок первичноротых уже был сегментированным, его тело делилось на парасегменты. У кольчецов всё так и осталось, а у членистоногих (и у других Ecdysozoa с сегментированным телом) на эту первичную сегментацию наложилась последующая перекройка сегментов, происходящая на более поздних стадиях эмбрионального развития.
А что же с конечностями? У кольчецов всё просто: каждый парасегмент отращивает по паре конечностей (параподий). Каждая конечность состоит из клеток, происходящих только от одного парасегмента. У членистоногих, как выяснилось, конечности имеют такой же смешанный состав, как и сегменты взрослых животных: передняя и задняя части каждой конечности образуются из разных парасегментов. Казалось бы, из этого следует, что конечности кольчецов и членистоногих негомологичны. Иными словами, общий предок первичноротых, хоть и был сегментированным, конечностей скорее всего не имел; кольчецы и членистоногие приобрели свои парные придатки независимо друг от друга.
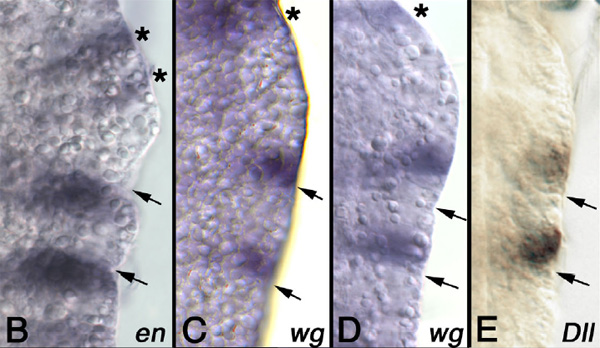
Для проверки этого предположения необходимо было очень детально изучить самые ранние этапы закладки конечностей в эмбриональном развитии членистоногих. Именно это и сделал специалист по сравнительной эмбриологии Никола-Михаэль Прпич (Nikola-Michael Prpic) из Гёттингенского университета (Georg-August-Universitat Gottingen). Он исследовал экспрессию гомеобоксных генов (Homeobox) engrailed (en), wingless (wg) и Distal-less (Dll) у личинки одного из самых примитивных членистоногих — рачка артемии. Гены en и wg управляют формированием парасегментов, причем en экспрессируется на переднем краю каждого парасегмента, а wg — на заднем. Это позволило исследователю точно локализовать границы парасегментов, которые иначе практически невозможно разглядеть.
|
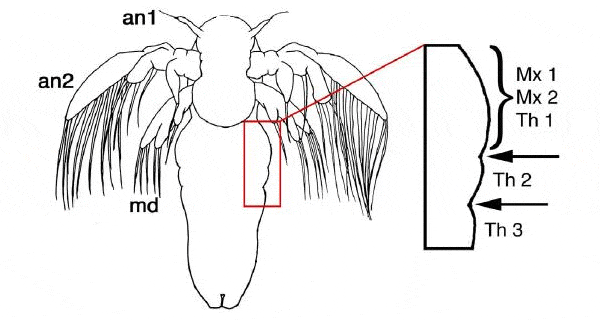
Ген Dll управляет развитием конечностей. Начинается всё с одной-единственной клетки, в которой ген Dll вдруг становится активным (начинается производство регуляторного белка, кодируемого этим геном). Затем экспрессия гена Dll распространяется на соседние клетки, из которых впоследствии образуется конечность. Таким образом, задолго до того, как зачаток конечности становится видимым, его можно обнаружить по активности гена Dll. Именно это и сделал Прпич. Ему удалось локализовать зачатки первой и второй пар грудных конечностей на очень ранней стадии их развития, когда эти зачатки представляют собой всего лишь небольшие группы клеток с включившимся геном Dll.
Оказалось, что зачаток конечности поначалу не выходит за пределы «своего» парасегмента. Иными словами, каждый зачаток состоит из клеток только одного парасегмента — точно так же, как у кольчатых червей. Позже область экспрессии гена Dll расширяется, и тогда уже в состав зачатка начинают входить и клетки другого (заднего) парасегмента.
|
Полученный результат — весомый довод в пользу того, что конечности у кольчецов и членистоногих все-таки имеют единое происхождение (то есть гомологичны друг другу). В обоих типах животных они начинают формироваться в соответствии с первичной сегментацией: каждый парасегмент отращивает по паре конечностей. Прпич вполне резонно предполагает, что так же обстояло дело и у общего предка первичноротых. Кольчецы сохранили этот исходный план развития, а у членистоногих на него наложились две позднейших модификации: 1) пересегментация, 2) расширение зачатка конечности, в результате чего в состав зачатка стали входить клетки заднего парасегмента.
|
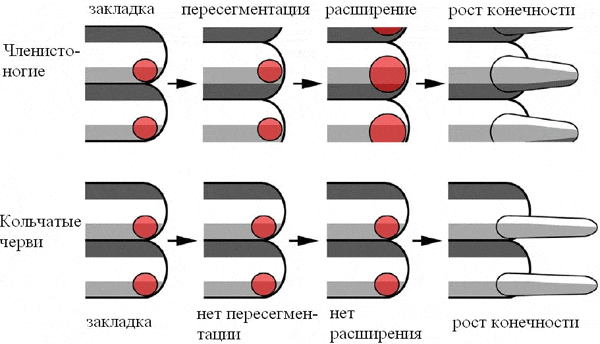
Если конечности у кольчецов и членистоногих гомологичны, значит общий предок этих двух типов животных (который «по совместительству» был также и общим предком первичноротых) тоже должен был обладать парными конечностями. Сегментированное животное с парными конечностями просто обязано было иметь также и вторичную полость тела — целом (целом служит червям чем-то вроде гидравлического скелета — наполненные жидкостью целомические пузыри необходимы как опорные образования для движения при помощи парных конечностей). Получается, что общий предок первичноротых был очень похож на типичного кольчатого червя. А если так, то старая классическая идея о происхождении членистоногих от кольчатых червей оказывается не такой уж и ошибочной.
Если результаты Прпича подтвердятся и будут признаны научным сообществом, то придется признать, что некоторые группы первичноротых эволюционировали по пути дегенерации (упрощения). Их предок был сегментированным животным с парными конечностями и целомом, однако многие его потомки впоследствии утратили сегментацию и конечности (моллюски, брахиоподы), а некоторые потеряли также и целом (немертины, плоские и круглые черви).
Источник: Nikola-Michael Prpic. Parasegmental appendage allocation in annelids and arthropods and the homology of parapodia and arthropodia // Frontiers in Zoology. 2008. V. 5. P. 17.
![]()