![]()
Риск вымирания редких видов может оказаться больше, чем мы думаем
|
Исследование американских биологов показывает, что принятые сейчас математические модели популяционной динамики могут сильно ошибаться в отношении небольших популяций. Возможно, потребуется пересмотр риска вымирания малочисленных видов.
Из наблюдений за живой природой известно, что численность какого-то вида на той или иной территории обычно не постоянна, а меняется из года в год. Очевидно, что если из-за этих колебаний популяция в какой-то момент резко сократится, то она уже не восстановится, а просто вымрет. Это становится особенно важным для малочисленных видов — у них вымирание популяции может означать исчезновение вида с лица Земли.
Один из главных возникающих здесь вопросов — в чём причина колебаний численности и можно ли как-то на нее повлиять? Для того чтобы на этот вопрос ответить, нужны не только полевые наблюдения, но и адекватные математические модели, которые помогли бы понять причины этих колебаний. Такие модели популяционной динамики, конечно, разрабатываются давно, и многие из них уже десятилетиями входят в учебники по математическому моделированию в экологии.
В последнем выпуске журнала Nature появилась статья, в которой утверждается, что все использовавшиеся до сих пор модели сильно ошибаются, когда речь заходит о малочисленных популяциях. В этих моделях, говорят авторы, не в полной мере учитываются факторы случайности при появлении на свет и развитии отдельных особей. Построенная авторами новая, более сложная модель, учитывающая эти факторы, приводит к резкому увеличению риска вымирания редких видов.
Чтобы изложить ситуацию подробнее, начнем с двух выводов, к которым пришли экологи после многочисленных попыток адекватно, без вычислительных артефактов, описать колебания реальных популяций:
- численность популяции требуется рассчитывать дискретными шагами по времени, а не плавной эволюцией (говоря на языке математики, надо использовать не дифференциальные, а конечно-разностные уравнения);
- нельзя пренебрегать случайными вариациями (то есть формулы должны содержать не только детерминированные, но и случайные слагаемые).
Одна из классических моделей, используемых для моделирования популяционной динамики, — модель Рикера (Ricker_model) — основывается на такой формуле для расчета численности популяции в i+1-ом поколении по ее значению в предыдущем, i-ом поколении:
Ni+1 = R Ni e–?Ni
Коэффициент R > 1 отражает среднюю плодовитость в расчете на одну особь, а коэффициент ? (много меньше 1) характеризует тот порог, начиная с которого популяция начинает вымирать от перенаселения: если в каком-то поколении количество особей заметно превышает 1/?, то в следующем поколении от популяции мало что останется.
Это — детерминистическая модель; в ней начальное количество особей полностью и однозначно определяет всю последующую эволюцию. В современных моделях в нее добавляют элемент случайности, то есть считают, что величина Ni+1 не строго задается этой формулой, а может слегка от нее отличаться в ту или другую сторону.
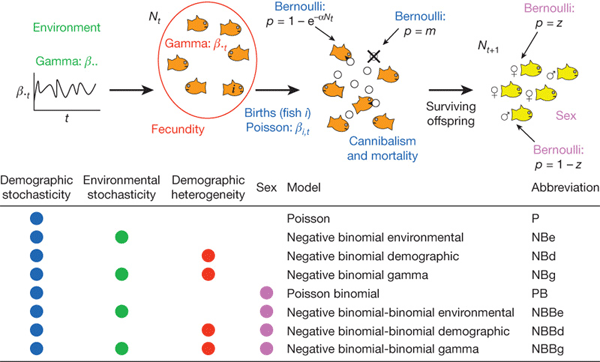
Авторы новой работы модернизировали эту классическую модель Рикера. В отличие от предыдущих моделей, они работали не на уровне всей популяции в целом, а на уровне индивидуальных особей. При этом они учитывали сразу четыре источника случайностей:
1) колебания внешних условий (климат, пища, естественные враги);
2) демографическую случайность (каждая особь имеет независимую вероятность выжить или умереть);
3) случайность в определении пола особи;
4) демографическое разнообразие (то есть особи имеют вовсе не одинаковую способность к выживанию и репродукции).
Авторы построили несколько математических моделей, включавших те или иные факторы из этого списка (см. рис. 1).
В рамках всех этих моделей была изучена динамика небольшой популяции (численностью в несколько десятков особей) при различных значениях параметров. Более конкретно, путем многократного прогона моделирования определялось среднее время жизни популяции, то есть количество поколений до ее полного вымирания.
На рис. 2 показаны типичные результаты такого анализа. Здесь по вертикальной оси показано среднее время жизни популяции, которая вначале насчитывала 30 особей, а по горизонтальной — значение коэффициента воспроизводства R. Разные кривые отвечают разным моделям.
|
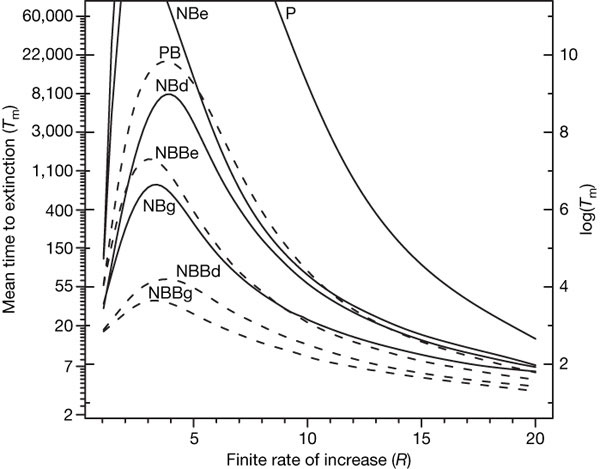
Два важных свойства этих результатов бросаются в глаза.
Во-первых, видно, что самая простая модель, отмеченная буквой P, проходит на графике выше всех. Например, при R = 10 и при неизменных внешних условиях начальная популяция из 30 особей сможет продержаться, согласно этой модели, несколько тысяч поколений. Однако если при тех же постоянных условиях учесть случайность в определении пола и демографическое разнообразие (кривая NBBd), то среднее время выживания падает до десятка поколений!
Во-вторых, изменчивость внешних условий перестает играть большую роль при переходе к более сложным моделям. Если различие между кривыми P (простая модель в постоянных условиях) и NBe (простая модель в изменчивых условиях) огромное, то разница между NBBd (сложная модель в постоянных условиях) и NBBg (сложная модель в изменчивых условиях) очень невелика.
Из этих результатов следуют вполне конкретные выводы. Опираясь на простейшие модели популяционной динамики, можно здорово ошибиться в оценке риска и причин вымирания малочисленных видов. Даже если поместить исчезающий вид в «естественные условия» и остановить его истребление, то всё равно имеется риск спонтанного исчезновения вида, причем намного больший, чем считалось до сих пор.
Может возникнуть вопрос: а почему ученые уверены, что более сложная модель точнее описывает реальную ситуацию? Для ответа на этот вопрос авторы статьи провели классический эксперимент. Они взяли 60 инкубаторов, поместили в них разное количество особей жука Tribolium castaneum, и дали им срок 24 часа для того, чтобы отложить яйца. Затем убрали взрослых особей, оставили инкубаторы на 34 дня для созревания яиц, а по истечении этого срока — сосчитали особей в новом поколении. Когда ученые попытались описать результаты этого опыта с помощью всех моделей, то оказалось, что точнее всего результаты описывает самая сложная модель.
В заключение стоит, однако, подчеркнуть, что для более глубокого понимания динамики реальных популяций, особенно на масштабе многих поколений, требуется включать в рассмотрение и генетические механизмы регуляции численности и состава популяции, социальное поведение животных и другие факторы. Это может привести к ситуации, когда параметры, считавшиеся константами, начинают изменяться. Без детального исследования трудно сказать, изменится ли при этом характер эволюции.
Источник: Brett A. Melbourne, Alan Hastings. Extinction risk depends strongly on factors contributing to stochasticity // Nature. V. 454. P. 100–103 (3 July 2008).
![]()