![]()
В поисках универсального закона устройства биологических сообществ, или Почему экологи потерпели неудачу?
|

В любом природном сообществе (будь то планктон озера, деревья тропического леса или совокупность птиц, обитающих на определенной территории) всегда можно найти несколько видов массовых, доминирующих по численности и гораздо большее число видов редких, малочисленных. Если у нас имеется большая (в несколько тысяч особей) проба из сообщества, то, разбив все обнаруженные в ней виды на классы обилия и определив число видов в каждом классе, мы можем построить гистограмму распределения частот встречаемости видов разной численности. График, проведенный по вершинам столбиков гистограммы, представляет собой сильно вогнутую кривую, напоминающую гиперболу. Подобный результат, выявляемый при анализе практически любого сообщества, на самом деле не очевиден и нуждается в объяснении. Кажется, более естественным было бы ожидать преобладания в природе видов, имеющих какую-то среднюю численность, при гораздо меньшем числе видов редких или, наоборот, массовых. Хотя проблема изучается уже более полувека, удовлетворительного объяснения наблюдаемой картины соотношения численностей разных видов пока нет. По мнению Брайена Макгилла (Brian J. McGill) c биологического факультета Университета Макгилла (Монреаль, Канада) и его соавторов, ситуацию, сложившуюся в изучении структуры сообществ, можно охарактеризовать как «коллективную неудачу» (collective failure). Выводы этого исследования, опубликованные в журнале Ecology Letters, могут быть поучительными не только для неудачливых экологов, но и всех, кому приходится сталкиваться с отсутствием прогресса в решении той или иной научной проблемы.
|
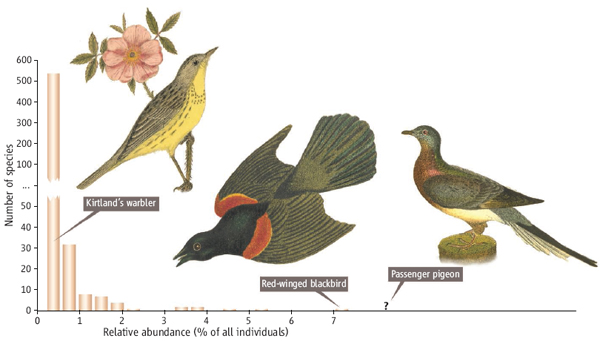
Выдающийся американский эколог Джордж Эвелин Хатчинсон (G. Evelyn Hutchinson, 1903–1991) одну из своих статей, вышедшую в 1959 году, назвал «Дань Святой Розалии, или Почему так много видов животных?» (Homage to Santa Rosalia, or why are there so many kinds of animals?, PDF, 30 Кб). В маленьком прудике около церкви Св. Розалии на горе Монте Пеллегрино около Палермо (Сицилия) он обнаружил двух водяных клопов одного рода, но различающихся размерами. Рассуждения о том, как виды могут сосуществовать несмотря на конкуренцию, привели Хатчинсона к постановке важных вопросов о том, что определяет видовую структуру сообществ. Однако проблема соотношения численностей разных видов в сообществе имеет на самом деле куда более давнюю историю.
|

Так, еще в 1932 году появилась работа японского исследователя Мотомуры, предложившего модель, согласно которой виды одного сообщества, ранжированные по убыванию численности, используют одну и ту же долю оставшегося общего ресурса. Например, если самый многочисленный вид потребляет 70% ресурса (и соответственно, такова его численность), то второй вид потребляет 70% того, что осталось от первого, следующий по убыванию численности вид — 70% от того, что осталось после первого и второго, и так далее. Через 25 лет американский эколог, ученик Хатчинсона Роберт Макартур (Robert MacArthur), используя сходную логику, предложил свою «модель разломанного стержня» (brocken stick model), в которой лимитирующий ресурс (одномерное пространство ниши) моделировался стержнем, случайным образом разломанным в разных местах. Численности видов были пропорциональны длинам образовавшихся отрезков. Хотя модель Макартура удовлетворительно описывала соотношения численностей разных видов птиц, сам автор позднее признал ее в теоретическом отношении несостоятельной.
|

Совершенно другой подход к решению проблемы видовой структуры сообщества использовал известный специалист по математической статистике Рональд Фишер (Ronald Fisher). Работая на Ротамстедской экспериментальной станции (Rothamsted Experimental Station) в Англии, он помог энтомологам А. Корбету и К. Уильямсу статистически обработать результаты отлова ночных бабочек с помощью световых ловушек. Фишер предположил, что распределение числа видов бабочек, имеющих разную численность, лучше всего описывается логарифмическими сериями (Fisher, Corbet, Williams, 1943). Максимальное число видов, согласно модели Фишера, приходится на категорию самых редких, представленных в пробах порой всего одним-единственным экземпляром. Чуть позднее появилась работа американского исследователя Ф. Престона, который использовал логарифмические классы обилия — «октавы», как он их называл. В первую октаву входили виды, представленные в выборке одной особью, во вторую — двумя особями, в третью — 3-4, в четвёртую — 5-8, в пятую — 9-16 и так далее (см. такую шкалу на рис. 4 и 5). Гистограмма соответствующего распределения напоминала классическую Гауссову кривую нормального распределения. Иными словами, распределение оказалось логнормальным: самых редких видов было всё же меньше, чем слегка более многочисленных. Престон полагал, что все достаточно большие выборки из сообщества должны описываться логнормальным распределением. Если же проба недостаточна репрезентативна, то левая часть кривой нормального распределения («хвост» редких видов) оказывается отрезанной.
|
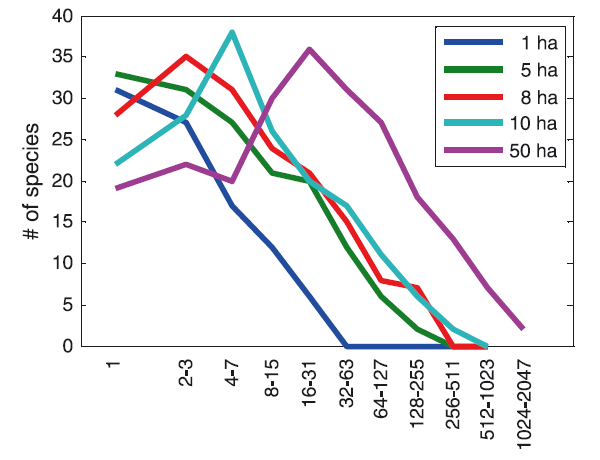
Макгилл и его соавторы (17 исследователей из 7 стран), проанализировав огромный массив литературы по проблеме видовой структуры сообществ, выявили более сорока моделей, описывающих соотношение численностей разных видов. Эти модели различались принципами устройства. Одни сводились к поиску математического распределения, которое бы удовлетворительно описывало некий набор эмпирических данных. Другие исходили из того, что особи размножаются, и, соответственно, можно использовать модели, основанные на теории ветвящихся процессов (branching processes). Третьи апеллировали к представлениям о динамике численности популяций. А были и такие, которые основывались на пространственном распределении видов или на том, как может делиться общий, ограничивающий всех ресурс (примеры последней категории — модели Мотомуры и Макартура).
|
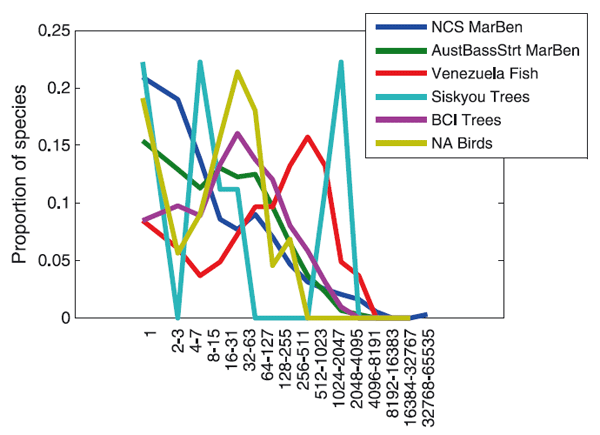
Однако, несмотря на столь большое число разных моделей, все они более или менее удовлетворительно демонстрировали один и тот же результат, а именно — «вогнутую кривую» (то есть распределение, характеризующееся множеством редких видов и очень малым числом видов массовых). Вряд ли такой результат можно считать существенным достижением, особенно учитывая ту огромную работу, которая была проделана множеством экологов уже не одного поколения. Скорее — это «коллективная неудача» (collective failure), причина которой, по мнению авторов обсуждаемой статьи, в том, что не были соблюдены методические установки, обеспечивающие научный прогресс. Совокупность таких установок, или «правил», Джон Платт (Platt, 1964) в свое время назвал «строгим выводом» (Strong inference). Апеллируя к этой старой, но вовсе не утерявшей своего значения работе Платта, Макгилл и его соавторы формулируют своего рода уроки прошлого — несколько ключевых положений, которые обязательно должны быть учтены, если мы хотим добиться прогресса в познании видовой структуры сообществ:
(1) Конкуренция гипотез. Ситуация была таковой, что новые гипотезы предлагались, а старые не отвергались. Модели не сравнивались между собой по результатам, то есть по тому, насколько хорошо они описывают реальность. Конкуренция гипотез отсутствовала.
(2) Множественные механизмы. Экологи порой забывали, что сходство паттернов (выявляемых регулярностей) вовсе не означает обязательного сходства процессов, которые могут их порождать. Да и в математике одна структура может быть получена разными путями.
(3) Решающие свидетельства. Нередко предсказания разных моделей столь мало между собой различались, что сравнивать их было очень сложно. Если какая-то модель удовлетворительно объясняет 99,1% вариации реальных данных, а другая только 99,0% — можем ли мы утверждать, что первая модель действительно лучше? Ведь статистическая ошибка тех данных по природным сообществам, которые имеются в нашем распоряжении, весьма велика, значительно больше различий, даваемых разными моделями.
(4) Робастные оценки. Суждения о преимуществе той или иной модели должны основываться на робастных оценках, то есть устойчивых при использовании разных критериев.
(5) Применимость модели по отношению к разным множествам данных. Нередко определенная модель хорошо описывает одну группу данных, но плохо описывает другую. Конечно, можно думать, что механизмы, ответственные за формирование структуры сообщества деревьев, отличны от механизмов, формирующих сообщество рыб. Но когда речь идет об участках тропического леса на разных континентах, то правило бритвы Оккама («не следует множить сущее без необходимости») заставляет нас ожидать сходных процессов.
Несмотря на общий критический настрой обзора Макгилла и др., в нём есть и оптимистичные ноты. Универсальность «вогнутой кривой» и ее возможная связь с разными макроэкологическими процессами, проявляющимися на больших совокупностях разных видов, населяющих крупные территории и акватории, позволяет надеяться на то, что в конце концов будет сформулирована некая объединяющая экологическая теория. Авторы статьи подчеркивают, что от теории в экологии не следует ожидать прогнозов, возможных в физике (например, таких, как столь любимый известным методологом науки Имре Лакатосом пример предсказания существования Нептуна по отклонениям орбиты Урана). Они разделяют мнение Розенцвейга и Абрамского (Rosenzweig, Abramsky, 1997), которые полагают, что не только теория видовой структуры сообщества, но и вся экология продвигаются вперед посредством не радикальной смены парадигм, а, скорее, методом, который они окрестили «dipswitch testing».
DIP-переключатель (Dip switch) — это набор связанных переключателей, которым еще недавно пользовались при настройке компьютеров (DIP, dual-in-line package — корпус с двухсторонним расположением выводов). Фактически, логика подхода «dipswitch testing» сводится к тому, что экологи могут давать множество бинарных предсказаний типа «Y возрастает, если возрастает X». Одно такое бинарное предсказание с вероятностью 50 на 50, конечно, очень слабое, но если какая-то модель (гипотеза) дает множество таких качественных (но проверяемых) предсказаний, то мы вправе считать данную модель хорошей.
Макгилл и его соавторы призывают экологов рассматривать проблему структуры сообщества не изолированно, а в более общем контексте, в частности изучать то, как влияют на структуру абиотические факторы, как проявляется структура на разных трофических уровнях или как на ней сказывается распространение заносных, «инвазивных видов» (см.: Invasive species), часто становящихся необычайно массовыми.
Источник: Brian J. McGill, Rampal S. Etienne, John S. Gray et al. Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework (полный текст статьи в открытом доступе) // Ecology Letters. 2007. V. 10. No. 10. P. 995–1015.
![]()