![]()
Взлет и падение видов: новые данные подтверждают старую идею «эволюционного цикла»
|
Детальный анализ пространственно-временного распространения 140 вымерших видов моллюсков показал, что виды проходят в своем развитии этапы роста, расцвета и упадка. После появления вида его «распространенность» (площадь ареала и частота встречаемости) постепенно растет в течение нескольких миллионов лет, ненадолго достигает максимума и затем постепенно снижается. Виды редко вымирают внезапно, находясь на пике численности; вымиранию обычно предшествует длительный период упадка. Новые данные подтвердили старую идею «эволюционного цикла», популярную в первой половине XX века, но затем вышедшую из моды.
Идея о том, что таксоны (виды, роды, семейства и т. д.) проходят в своем развитии закономерные стадии молодости, зрелости и старости, высказывалась многократно начиная еще с XIX века. В принципе, такая этапность в развитии многих крупных таксонов видна в палеонтологической летописи, что называется, невооруженным глазом. Объяснения этому предлагались самые разные: от довольно абстрактных и даже идеалистических (вроде исчерпания «жизненной силы») до вполне материалистических и конкретных (см. ссылки внизу). Во второй половине XX века идея эволюционного, или филогенетического цикла стала выходить из моды — отчасти из-за идеалистического «привкуса», отчасти из-за отсутствия надежных статистических доказательств реальности данного феномена. Чтобы получить доказательства, нужны были обширные, подробные и достоверные палеонтологические базы данных.
Сегодня такие базы уже существуют, и среди них одной из самых качественных и детализированных по праву считается FRED (The Fossil Record Electronic Database), содержащая сведения о палеонтологических коллекциях, собранных в Новой Зеландии и прилегающих районах.
Майкл Фут (Michael Foote), известный специалист по эволюции биоразнообразия из Чикагского университета, и его коллеги из Новой Зеландии использовали эту базу данных для поиска закономерностей в развитии разных видов и родов ископаемых новозеландских моллюсков. Надо сказать, что большинство имеющихся палеонтологических сводок позволяет проводить статистический анализ только на родовом и более высоких таксономических уровнях, а данные на уровне видов обычно либо недостаточно детальны, либо сомнительны. FRED в этом смысле представляет собой приятное исключение.
Исследователи выбрали для своего анализа 140 видов кайнозойских моллюсков, данные по которым оказались наиболее достоверными и подробными. Все эти виды появились и вымерли примерно от 40 до 3 млн лет назад. Были использованы только вымершие виды, чтобы можно было проследить всю их судьбу целиком — от рождения до смерти. Для датировки коллекций в FRED используется ярусная (вековая) шкала, то есть возраст каждой коллекции (и, соответственно, каждой отдельной находки) известен с точностью до века (это в среднем примерно 2 млн лет). Чтобы при таком временном разрешении можно было заметить какие-то изменения, происходящие с видом во времени, ученые рассматривали только виды, известные не менее чем из трех последовательных веков. Все исследованные виды, кроме двух, являются новозеландскими эндемиками, то есть не известны из других регионов.
Для каждого вида была вычислена его «встречаемость» или «распространенность» (occupancy) на каждом этапе его существования, то есть отдельно для каждого века, в котором существовал вид. В качестве меры «встречаемости» использовали просто-напросто долю коллекций данного возраста, в которых присутствует данный вид. Авторы показали, что эта мера коррелирует с площадью видового ареала (что вполне естественно). Кроме того, сравнивая коллекции современных и ископаемых новозеландских моллюсков и «встречаемость» в них одних и тех же видов, исследователи показали, что палеонтологическая «встречаемость» очень хорошо коррелирует с современной. Это значит, что результаты исследования не должны быть сильно искажены неполнотой летописи.
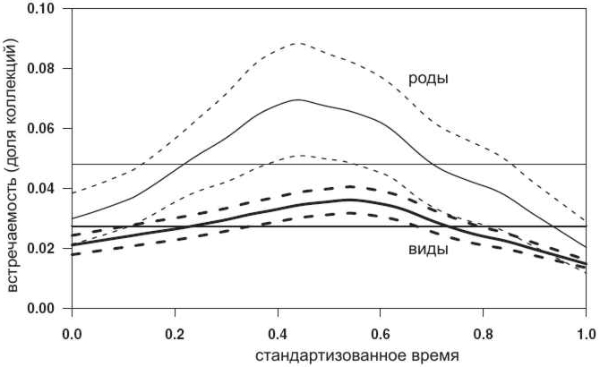
Полученные результаты показаны на рисунке. Оказалось, что 56% видов демонстрируют схожую динамику встречаемости: в начале своей истории они сравнительно редки, потом в какой-то момент достигают максимальной встречаемости, а затем снова становятся более редкими. Иными словами, имеется один выраженный кратковременный пик встречаемости, не совпадающий ни с первым, ни с последним веком существования вида. Диаграммы, соответствующие этим видам, на рисунке выделены черным. Лишь 16% видов достигли максимального распространения в самый первый век своей жизни, 14% — в последний, а по остальным 14% видов результаты получились неопределенные (либо вообще нет выраженного пика, либо их два и т. п.).
Затем авторы объединили все 140 диаграмм в один обобщенный график, показывающий усредненную динамику развития вида; аналогичная работа была проделана и на уровне родов (см. рисунок в начале заметки). Получился самый типичный «эволюционный цикл» с ростом, коротким расцветом и упадком, который для изученных 140 видов неизменно заканчивался вымиранием. На родовом уровне картина получилась точно такая же. Впрочем, на родовом и семейственном уровнях такие картинки были получены и раньше (в качестве меры «встречаемости», за неимением данных по отдельным находкам, использовалось видовое или родовое разнообразие таксона).
Например, для семейств морских ежей при помощи сходной методики нами был получен очень похожий куполообразный усредненный график еще в 1994 году (Марков А.В., Наймарк Е.Б. 1994. Динамика филогенетических процессов в надвидовых биосистемах // Палеонтол. журн. № 3 C. 3–12). Однако то, что на видовом уровне дело обстоит точно так же — это результат действительно новый и важный. Надо сказать, что те механизмы, которые предлагались ранее для объяснения «филогенетических циклов» у надвидовых таксонов (см. ссылки внизу), не подходят для объяснения такой же динамики на уровне видов, или, по крайней мере, нуждаются в серьезной модификации.
К статье прилагаются пространные дополнительные материалы с множеством графиков, которые доказывают достоверность и статистическую значимость полученных результатов.
|

Результаты, полученные Футом и его коллегами, не подтвердили предположение о том, что виды достигают максимального расцвета (наибольшего географического распространения, численности, биомассы и т. д.) вскоре после своего появления, то есть еще в «ранней молодости». Такое мнение было основано, в частности, на некоторых современных наблюдениях, когда под воздействием человека отдельные виды (например, случайно занесенные из другого региона) стремительно распространялись по огромным территориям. В природе, по-видимому, такое происходит редко, и новому виду обычно требуется несколько миллионов лет, чтобы полностью реализовать свой эволюционно-экологический потенциал, то есть реально заселить все места, которые он в принципе способен заселить.
Другой важный вывод состоит в том, что виды редко вымирают внезапно, находясь на пике своего развития. Вымиранию обычно предшествует довольно долгий период упадка. Это значит, что палеонтологическая история вида позволяет судить о вероятности его вымирания в наши дни: наибольшей опасности подвергаются те виды, которые уже миновали пик своего развития и находятся в фазе упадка.
Полученные данные противоречат также распространенному мнению о том, что в эволюции должны чередоваться короткие периоды «становления» и долгие периоды «стазиса». В действительности виды, по-видимому, почти не задерживаются на максимальном достигнутом уровне и практически сразу переходят от роста к упадку.
Характерно, что авторы даже не пытаются предложить какую-то связную гипотезу о механизмах загадочного «старения» видов. Для подобных гипотез пока нет достаточных фактических оснований, а голословные рассуждения на страницах Science не приветствуются.
Источник: Michael Foote, James S. Crampton, Alan G. Beu, Bruce A. Marshall, Roger A. Cooper, Phillip A. Maxwell, Iain Matcham. Rise and Fall of Species Occupancy in Cenozoic Fossil Mollusks // Science. 2007. V. 318. P. 1131–1134.
![]()