![]()
Разгадан механизм регуляции роста коренных зубов
|

Финские ученые разгадали механизм регуляции роста коренных зубов млекопитающих. Оказалось, что темпы роста и размер моляров зависят от количества белка-активатора, который выделяется клетками челюсти, и белка-ингибитора, который выделяется растущим зубом. Таким образом, каждый растущий зуб определяет размер последующего. Это удалось подтвердить экспериментами и данными по соотношению зубов у мышиных грызунов.
Примерно год назад появилось сообщение о том, что японским ученым удалось вырастить в пробирке зуб и потом вживить его в челюсть 8-недельному мышонку (см. Takashi Tsuji et al. The development of a bioengineered organ germ method // Nature Methods. 2007. V. 4. P. 227–230). Зуб отлично прирос, то есть в нем сформировались кровеносные сосуды, нервы, необходимые для функционирования нормального живого зуба. Всем ясно, какие перспективы у этого эксперимента. Он показывает принципиальную возможность замены потерянного зуба на его выращенный в пробирке аналог.
Но, естественно, от принципиальной возможности до практического исполнения в рядовой клинике лежит долгий научный путь. Один из ключевых вопросов — как управлять процессом формирования зуба. Для этого нужно в деталях понять, как регулирует рост зуба сам организм млекопитающего. Известно, что развитием зуба управляет целый блок генов, которые экспрессируются и в челюсти, и в самом зубе. Однако генные механизмы регуляции одонтогенеза (то есть развития зубов) пока в точности не известны. Кроме того, ученые не склонны всю регуляторику сваливать на гены.
Группа финских ученых, работающих в Институте биотехнологии Хельсинкского университета и на факультете экологии и эволюции Университета штата Нью-Йорк в Стони-Бруке (США), экспериментально вывела алгоритм регуляции развития коренных зубов, который работает у всех грызунов (ученые предполагают, что вообще у всех млекопитающих).
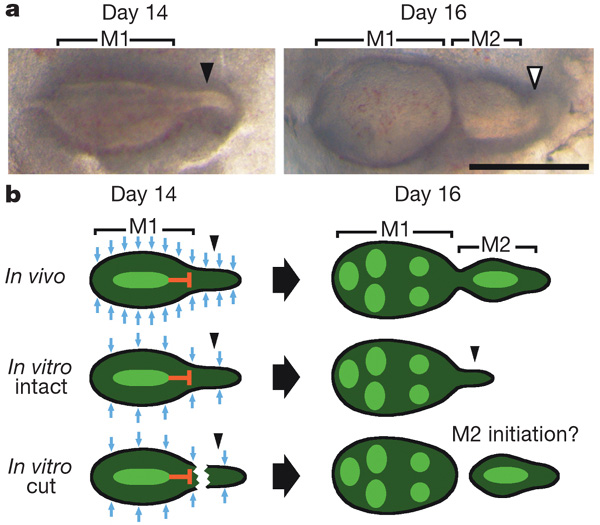
У млекопитающих, за редким исключением, имеется по три коренных зуба справа и слева в верхней и нижней челюсти. Они развиваются всегда от центра к периферии. Сначала прорезается первый коренной зуб, затем мезенхимные клетки челюсти посылают сигнал о начале роста второго коренного зуба. И тогда от первого коренного зуба отпочковывается зачаток второго коренного зуба. Одновременно первый моляр вырабатывает биохимические факторы, тормозящие развитие второго зуба.
Таким образом, развитие второго зуба контролируется двумя противодействующим силами: силой активирующего сигнала от мезенхимы челюсти и силой тормозящего сигнала от растущего первого моляра. От их соотношения и будет зависеть размер второго зуба. На определенной стадии развития второго моляра от него отрастает назад зачаток третьего моляра. Сроки закладки этого зачатка и размер третьего моляра опять-таки зависят от силы мезенхимного активатора и тормозящего сигнала, посылаемого вторым моляром. То есть темпы роста коренных зубов и их размеры зависят от разницы между количеством активатора и ингибитора. (Косвенно об этом свидетельствует и такой факт: мутации, блокирующие фактор-ингибитор, приводят к образованию дополнительных моляров или дополнительных бугорков на коронках.)
Развивающийся зуб определяет судьбу следующего за ним зуба, а тот, в свою очередь контролирует судьбу следующего. И не требуется никаких особых наборов генов на каждый зуб. Теоретически, для регуляции развития всех моляров достаточно двух генов: один отвечает за количество мезенхимного активатора, а другой включает выработку ингибитора в развивающемся зубе. В действительности, при росте зубов работает не один такой ингибитор, а несколько, например эктодин (Ectodin) и фоллистатин (Follistatin).
Чтобы доказать этот гипотетический каскадный механизм регуляции формирования моляров, были выполнены остроумные эксперименты. Суть их состоит в следующем. Зачатки эмалевого органа мышей — зачаточная ткань, из которой развиваются зубы у млекопитающих, — выращивали в культуре (см. обзор о культивировании зубов млекопитающих «Зубы из пробирки»). Зубные зачатки первого моляра, как и положено, отпочковали второй моляр. В этот момент зачаток второго моляра отделили от первого, и он дальше развивался самостоятельно. Осталось сравнить скорости развития второго моляра у неотделенных и отделенных вторых моляров и сроки закладки третьих моляров.
По идее, отделенные вторые моляры должны расти быстрее, потому что на них не действуют ингибиторы первого моляра. Как и предполагалось, отделенные зачатки вторых моляров развивались быстрее и становились больше, чем те, что находились в паре с первым моляром. Также у них и закладка третьих моляров происходила на несколько дней раньше, чем у пары первый—второй моляр. Третьи моляры, спаренные с отделенными вторыми молярами, тоже достигали сравнительно больших размеров, чем их аналоги в нормальной триаде моляров. Ясно, что как только устранили тормозящий эффект первого моляра, рост значительно ускорился.
|
Ученые решили проверить, насколько приложима эта простая каскадная схема регуляция к другим представителям мышиных. Ведь если работает правило сложения активации-ингибирования, то соотношение размеров моляров не может быть случайными. Если второй моляр больше первого, то это означает, что ингибитора немного, и третий моляр будет еще больше второго. А если первый моляр большой, а второй маленький, то это означает, что ингибирование усилено, и третий моляр должен быть совсем маленький. В крайнем случае третий моляр не вырастает вовсе. Действительно, у представителей мышиных соотношение моляров хорошо укладывается в эту модель. А у бобровой крысы (Hydromys chrysogaster) имеется упомянутый крайний вариант: у нее очень большие первые моляры, маленькие вторые моляры, а третьих вообще нет.
Каскадная модель регуляции роста моляров объясняет одну известную, но доселе загадочную закономерность: у всех приматов второй моляр всегда занимает ровно треть молярного пространства челюсти. Относительный размер второго моляра (а — количество активатора, и — количество ингибитора) равен:
М2 / (М1 + М2 + М3) = (а/и) / [1 + а/и + (2а/и – 1)] = 1/3
Так что модель предсказывает, что относительный размер второго моляра постоянен не только у приматов, но и у всех других млекопитающих.
Несмотря на очевидную практическую значимость открытия каскадной регуляции развития зубов, авторы склонны подчеркивать скорее теоретическое значение своего открытия. Их рассуждения уводят нас в область механизмов эволюции. Эволюция распределила множество видов грызунов по различным экологическим нишам: тут и питание различной пищей, и разные местообитания. В соответствии с типами питания у них различаются и зубы.
Однако, как выяснилось, развитие и эволюцию зубов определяют не случайные мутации, «подправляющие» их форму и размер. Форму и размер зубов (читай — тип пищи) определяет жестко заданное в онтогенезе соотношение активатора и ингибитора. Так что эволюция группы идет в пределах некоторого заранее заданного диапазона разрешенных онтогенезом вариантов. При этом добавление к каскадной регуляции еще одного переключателя может перевести онтогенез на другое направление, и это будет означать появление нового большого таксона.
Идея о том, что закономерности онтогенеза канализируют (направляют) эволюцию, резко ограничивая число возможных эволюционных преобразований, развивается в фундаментальном труде И. И. Шмальгаузена Организм как целое в индивидуальном и историческом развитии (см. также: Эпигенетическая теория эволюции).
Источник: K. D. Kavanagh, A. R. Evans, J. Jernvall. Predicting evolutionary patterns of mammalian teeth from development // Nature, 2007. V. 449. P. 427–433.
Елена Наймарк
![]()
Златобрюхая бобровая крыса — крупный грызун длиной 20-35 см и до 1,5 кг весом с перепончатыми лапами, обитает в болотах, реках, озерах и морских побережьях, питается лягушками, моллюсками, рыбами, может утащить и яйца уток, и даже птенца. У них кроме резцов имеется крупный первый коренной зуб и маленький второй. А третьего моляра нет вовсе: он не развивается из-за большого количества ингибиторного белка, выделяемого растущими первым и вторым моляром. (Рис. с сайта commons.wiki.dorotka.info)