![]()
Конкурирующие виды могут сосуществовать и в одной, и в разных нишах
|

Согласно традиционным воззрениям, разные виды организмов сосуществуют постольку, поскольку различаются их экологические ниши. Но новая гипотеза нейтральности утверждает, что виды могут сосуществовать, если они очень сходны по своим демографическим характеристикам и фактически занимают одну нишу. Предложенная недавно простая модель позволяет примирить эти, казалось бы, несовместимые взгляды на природу экологических сообществ. Сосуществование конкурирующих видов в рамках этой модели может достигаться как при расхождении их экологических ниш, так и при их близости.
В течение долгого времени считалось, что виды организмов, входящих в одно сообщество (живущие вместе и зависящие от общих ресурсов), должны занимать разные экологические ниши. В противном случае нарушался бы принцип конкурентного исключения (принцип Вольтерры—Гаузе), и тот вид, который быстрее или эффективнее других использует дефицитный ресурс, вытеснил бы остальные. Однако известны были и такие сообщества — например, микроскопических водорослей, взвешенных в толще воды, или деревьев, произрастающих в одном лесу, — по отношению к которым предположить расхождение всех видов по разным нишам довольно трудно. И в том и в другом случае виды сосуществуют, хотя конкурируют за одни и те же ресурсы. Изучение таких сообществ приводило некоторых экологов к выводам, противоположным общепринятым.
Так, Стивен Хаббел (Stephen P. Hubbell), опираясь на свой опыт изучения тропических лесов, предложил «гипотезу нейтральности», согласно которой разные виды деревьев (а на одном гектаре тропического леса их может быть более сотни) сосуществуют благодаря своему сходству, а не различиям. При этом Хаббел и его сторонники вовсе не отрицают того, что разные виды в сообществе могут занимать разные ниши. Они только замечают, что есть и другой способ достичь стабильного сосуществования, а именно — быть максимально похожими по демографическим характеристикам, иметь сходные удельные (то есть в расчете на одну особь) скорости популяционного роста и скорости заселения освободившихся участков.
Гипотеза нейтральности вызвала огромный интерес среди экологов, расколов их сообщество на две партии — ее сторонников и ее противников. Найти убедительные эмпирические свидетельства в пользу той или иной гипотезы оказалось очень непросто. На фоне горячих дискуссий стали появляться работы и тех, кто ищет примирения. Одна из таких работ, опубликованная в последнем номере журнала Ecology Letters, так и называется: «Ниша для нейтральности». Её авторы, Питер Эдлер с Факультета природных ресурсов Университета штата Юта (США) и его коллеги из двух других университетов США, предложили модель, связывающую концепцию нейтрализма с традиционными представлениями о нишевой структуре сообщества. В основу модели положена идея Питера Чессона (Peter Chesson), рассматривающего нейтральность как крайний случай более общей теории, учитывающей прежде всего расхождение видов по разным нишам.
|
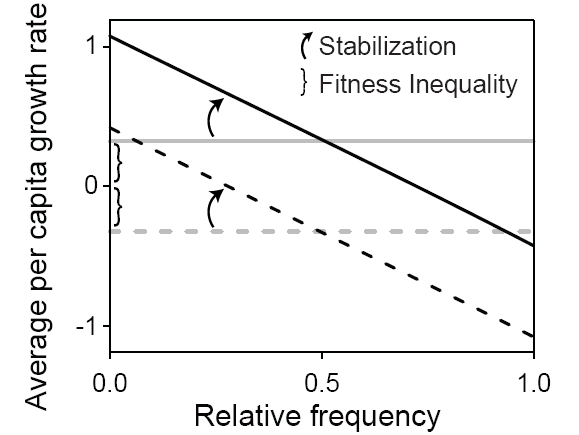
Эта модель предполагает, что в том случае, когда разные виды в сообществе занимают разные ниши, внутривидовая конкуренция должна иметь большее значение для ограничения численности вида, чем межвидовая. Соответственно, по мере того, как возрастает доминирование (точнее — относительная частота встречаемости) данного вида в сообществе, удельная скорость его популяционного роста должна падать (см. рис. 1). То, насколько быстро это происходит (то есть наклон линии относительно осей координат на рис. 1), трактуется как «степень стабилизации». Если же механизм разделения ниш не работает (гипотеза нейтральности), то удельная скорость популяционного роста остается постоянной независимо от относительной частоты встречаемости данного вида (линия идет параллельно оси Х на рис. 1). В этом случае необходимое условие сосуществования видов — близость скоростей их популяционного роста (на оси Y они должны располагаться по возможности ближе друг к другу).
|
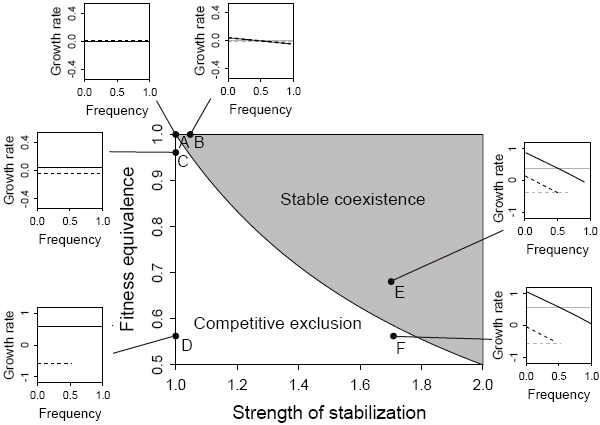
В обсуждаемой работе степень стабилизации (иными словами — степень расхождения видов по разным нишам) авторы сопоставили со степенью близости видов по демографическим характеристикам (рис. 2). Таким образом в одной модели (на поле одного графика) удалось объединить представление о нейтральности с традиционным взглядом на обязательное разделение ниш сосуществующих видов. Чем больше сходство видов по скорости их популяционного роста, тем менее значимую роль в поддержании их сосуществования играет «стабилизация» (то, насколько быстро снижается скорость популяционного роста вида при увеличении его относительной численности). И наоборот, чем сильнее различаются виды по скорости популяционного роста, тем большее значение приобретает «стабилизация».
Где на рис. 2 располагаются точки, соответствующие реальным сообществам, мы не знаем, но на основании этой модели авторы предлагают путь к количественной оценке тех параметров, которые и определят соотношение нейтральности и расхождения видов в каждом конкретном случае. Таким образом, Питеру Эдлеру и его коллегам удалось сделать реальный шаг на пути примирения диаметрально противоположных взглядов на природу сообщества.
Источник: Peter B. Adler, Janneke HilleRisLambers, Jonathan M. Levine. A niche for neutrality // Ecology Letters. 2007. V. 10 (2). P. 95–104 (полный текст: Pdf, 900 Кб).
![]()